Экспериментальный волновой иммунитет и волновая генетика. Теоретические модели.
Артюх В.Д., Гаряев П.П., Кокая А.А., Леонова-Гаряева Е.А., Мулдашев Э.Р., Мухина И.В., Смелов М.В., Тертышный Г.Г., Товмаш А.В., Чалкин С.Ф., Шатров Я.К., Ягужинский Л.С.[1]
|
|
Многие исследователи в своих работах указывают на опасность загрязнения гептилом окружающей среды, что негативно сказывается на здоровье человека и отрицательно влияет на экологию [http://www.seu.ru/conference/ecoprvo/geptil.htm]. В настоящем исследовании демонстрируется принципиальная возможность разработки технологии, позволяющей выработать у биосистем устойчивость к токсическому действию гептила с помощью электромагнитных полей. Теоретические основы для проведения такого рода исследований представлены на сайте http://www.wavegenetics.jino-net.ru/ . Прежде чем начать работу в этом направлении, нами был выполнен ряд первичных модельных экспериментов с использованием аллоксана. Аллоксан является цитотоксическим веществом с преимущественным поражением b-клеток поджелудочной железы и вызывающий сахарный диабет 1-го типа.
В предыдущей экспериментальной работе [Гаряев, Кокая и др., 2007] было отмечено, что модулированное поджелудочной железой и селезенкой широкополосное электромагнитное излучение (мШЭИ), генерированное гелий-неоновым лазером, влияет на течение экспериментального сахарного диабета у крыс, вызванного внутрибрюшинным введением аллоксана в дозе 200 мг/кг массы тела животного. Воздействие данным видом излучения приводило к увеличению продолжительности жизни животных в опытных группах по сравнению с контролем, нормализации уровня глюкозы в крови, способствовало регенерации ткани поджелудочной железы.
|
|
Цель настоящей работы – оценить эффект устойчивости животных к аллоксану при превентивном воздействии на них мШЭИ. Для этого был использован специальный лазер, имеющий взаимосвязанные комплементарные ортогональные поляризации пучка света. Генерацию ШЭИ проводили по схеме интерферометра Фабри-Перо, в которой рабочий лазерный луч многократно проходит через тонкие свежепрепарированные срезы (препараты) поджелудочной железы и селезенки здорового новорожденного крысёнка. Мы предполагаем, что препараты специфически модулируют лазерный луч определённым образом, вследствие чего данная система проявляет следующие способности: 1) усиливать ШЭИ, идущее из разрядного промежутка He-Ne лазера; 2) ШЭИ параметрически связано с препарат-модулированным лазерным лучом и вследствие этого приобретает высокую биологическую активность; 3) биологический эффект может наблюдаться на относительно больших расстояниях от источника ШЭИ; 4) осуществляется электромагнитная передача управляющей генетико-метаболической информации с биообьекта-донора на биообьект-реципиент. Переносчиком информации в этом случае выступает модулированное биообьектом-донором широкополосное электромагнитное поле (мШЭИ). В качестве биоструктур, зондируемых лазерным лучом в данной системе, могут быть использованы живые и/или квази-живые организмы, например, бактерии, вирусы, а так же переживающие органы и ткани, метаболиты и абиогенные вещества.
МЕТОДИКА ИССЛЕДОВАНИЯ. ФИЗИЧЕСКАЯ ЧАСТЬ.
Для получения модулированного биоструктурами широкополосного электромагнитного излучения (мШЭИ) применяли ранее разработанную нами биотехнологию работы с использованием гелий-неонового лазера [Гаряев, Тертышный, 1999]. Гелий-неоновый лазер мощностью 2 мВт и длиной волны 632.8 нм имеет две совмещенные, ортогональные линейно поляризованные моды излучения, одночастотные в каждой из них. Лазерный луч зондирует биоструктуры, т.е. свежевыделенные препараты поджелудочной железы или селезенки новорожденной крысы линии Wistar. Полупрозрачные препараты наносили на предметное стекло, накрывали покровным стеклом и помещали на оптической оси лазерный луч-препарат. Юстировку стекол с препаратами проводили таким образом, чтобы обеспечить частичное обратное отражение луча, модулированного препаратами, в резонатор лазера. Такой многопроходный режим позволяет препарату выступать в роли оптического коррелятора [Мазур, Грачев, 1985] и влиять на распределение вторичных мод излучения лазера. Оптические сигналы регистрировали и подавали на электронную схему, которая управляет режимом генерации лазера, при этом происходит частотная стабилизация когерентного излучения. В таком режиме работы лазер генерирует, помимо красного света, ШЭИ, модулированное препаратами, т.е. собственно мШЭИ. Расстояние от зондируемого препарата до активного элемента лазера 11см.
МЕТОДИКА ИССЛЕДОВАНИЯ. БИОЛОГИЧЕСКАЯ ЧАСТЬ.
В экспериментах использовали половозрелых самцов крыс линии Wistar в возрасте 5-6 месяцев, средней массой 180-220 г. Экспериментальный сахарный диабет вызывали путем внутрибрюшинного введения раствора аллоксана (200 мг/кг массы тела животных) после 24 часового голодания на фоне нормальных показателей уровня глюкозы в крови. Животные были разделены на 4 группы: 1-я группа контроль (n=20) - без воздействия ШЭИ; 2-я (n=20) и 3-я (n=20) группы – животные подвергались предварительному воздействию мШЭИ; 4-я группа (n=10) – плацебо, в которой ШЭИ не модулировалось биоструктурами, а лазерный луч проходил через пустые предметное и покровное стекла без срезов ПЖ и селезенки.
2-ю группу животных в момент воздействия располагали на расстоянии 20 м от источника мШЭИ в подвальном помещении лаборатории. Аллоксановый сахарный диабет в этой группе вызывали через месяц после последнего воздействия мШЭИ. 3-ю и 4-ю группы животных располагали на расстоянии 70 см от источника мШЭИ. Аллоксановый сахарный диабет в этих группах вызывали через сутки после последнего воздействия.
Воздействие мШЭИ на 2-ю и 3-ю группы проводили по 30 минут ежедневно, в течение 4-х дней по схеме: 10 минут воздействие мШЭИ, полученным в результате прохождения лазерного луча через препарат с тканью поджелудочной железы; 10 минут воздействие мШЭИ, полученным в результате прохождения лазерного луча через препарат с тканью селезёнки; 10 минут воздействие мШЭИ, полученным в результате прохождения лазерного луча через препарат с тканью поджелудочной железы.
На 4-ю группу (плацебо) оказывали воздействие ШЭИ, которое не модулировалось биоструктурами (лазерный луч проходил через пустые предметное и покровное стекла без срезов поджелудочной железы и селезёнки) по 30 минут ежедневно, в течение 4-х дней.
1-я группа (контроль) не подвергалась воздействию ШЭИ и мШЭИ.
Во время эксперимента оценивали общее состояние животных, фиксировали день гибели животных с момента введения аллоксана во всех наблюдаемых группах. Животных 2-ой и 3-ей групп наблюдали в течение 1,5 месяцев c момента введения аллоксана. У 8 крыс из 2-ой и 3-ей групп с максимальным подъемом уровня глюкозы в крови после введения аллоксана оценивали репродуктивную функцию (3 крысы из 2-ой группы и 5 крыс из 3-й группы).
Регистрацию уровня глюкозы в крови проводили глюкометром Ascensia Entrust фирмы Bayer. Диапазон измеряемого уровня глюкозы от 2,0 ммоль/л до 30,6 ммоль/л. Значения уровня глюкозы в крови выше 30,6 ммоль/л обозначали HI.
Изъятие тканей сердца, легких, печени, почек, селезенки и поджелудочной железы для макроскопического описания и гистологического исследования проводили: в контрольной группе на 3 и 4-е сутки с момента введения аллоксана, что соответствовало дню максимальной гибели животных; во 2-й и 3-й группах на 8-е сутки с момента введения аллоксана, а также на 42-е сутки эксперимента у животных после оценки репродуктивной функции самцов. Для гистологических исследований ткани фиксировали в 10% нейтральном формалине, обезвоживали в спиртах восходящей концентрации и заливали в парафин. Парафиновые срезы толщиной 5-7 мкм получали на микротоме Leica SM 2000R, окрашивали гематоксилином и эозином и анализировали с помощью микроскопа Leica DMLS. Видеоизображения получали на видеосистеме с помощью CCD-камеры.
Статистическая обработка результатов исследования проводилась с помощью статистических программных пакетов “Stastica 6.0”, MS-Exel” for Windows. Степень достоверности (р) определялась по критерию Стьюдента, с использованием доверительного коэффициента и числа степеней свободы (1) по таблице. Вычисление всех математических параметров проводилось по общеизвестным формулам на ПК.
Исследования в данной серии экспериментов показали, что применение указанной дозы аллоксана в контрольной и плацебо группах способствовало развитию сахарного диабета, осложненного токсическим повреждением ряда жизненно важных органов и систем. Это приводило к очень низкой выживаемости животных в группах. Напротив, во 2-ой и 3-ей группах наблюдалась устойчивость животных к действию аллоксана в разной степени выраженности.
В 1-ой группе (контроль) выживаемость животных после введения аллоксана на 2-е сутки составила 55%, а к 4-м суткам снизилась до 30% (Рис.1). Уровень глюкозы в крови у животных 1-ой группы на 2-е, 3-е и 4-е сутки достоверно отличается (p<0.05) от исходного значения (табл.1). Гибнущих животных 1-ой группы (контрольной) в терминальном состоянии подвергали эвтаназии (5 крыс), органы использовали для патоморфологического исследования. В контрольной группе отсутствовало самопроизвольное снижение уровня глюкозы в крови за период наблюдения (табл.1) Однако в этой группе была одна крыса, устойчивая к действию аллоксана, у которой уровень глюкозы в крови оставался в пределах физиологической нормы.
Таблица 1.Уровень глюкозы в крови у животных после введения аллоксана в дозе 200мг/кг массы
|
Группы |
Уровень глюкозы в крови в исследуемых группах, ммоль/л |
|||
|
Исходный день |
2-е сутки |
3-е сутки |
4-е сутки |
|
|
1-я группа (контроль), n=20 |
5,97±1,38 n=20 |
25,93±8,16** n=18 |
24,65±9,78** n=8 |
22,63±10,7** n=6 |
|
2-я группа (опыт), n=20 |
6,78±0,83 n=20 |
24,7±9,17*** n=20 |
18,99±8,0*** n=18 |
21,93±9,91*** n=18 |
|
3-я группа (опыт), n=20 |
5,23±0,69 n=20 |
8,00±6,32* n=20 |
6,44±4,39* n=20 |
4,88±2,90* n=20 |
|
4-я группа (плацебо), n=10 |
4,9±0,85 n=10 |
26,97±6,85**** n=8 |
_ ^ n=1 |
_ ^ n=1 |
* - уровень глюкозы в крови у животных 3-ей группы на 2-е, 3-е и 4-е сутки моделирования аллоксанового диабета достоверно отличается ( р<0.05) от уровня глюкозы в крови животных 1-ой и 2-ой групп на 2-е, 3-е и 4-е сутки, а так же достоверно отличается (р<0.05) от уровня глюкозы в крови у животных 4-ой группы на 2-е сутки;
** - уровень глюкозы в крови у животных 1-ой группы на 2-е, 3-е и 4-е сутки достоверно отличается
( p<0.05) от исходного значения;
***- уровень глюкозы в крови у животных 2-ой группы на 2-е, 3-е и 4-е сутки достоверно отличается
( p<0.05) от исходного значения;
****- уровень глюкозы в крови у животных 4-ой группы на 2-е сутки достоверно отличается ( p<0.05) от исходного значения;
^ - в 4-ой группе на 3-е и 4-е сутки наблюдения было одно выжившее животное;
Исходный день – день введения аллоксана.
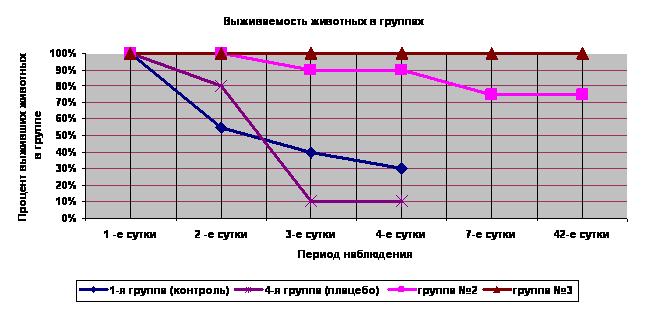
Рис. 1. Выживаемость животных (%) в исследуемых группах при моделировании аллоксанового диабета. Всем животным введён аллоксан в дозе 200 мг/кг массы. 1-я группа (контроль) воздействия ШЭИ и мШЭИ не проводилось. 2-я группа - проводилось воздействие мШЭИ, животные были расположены на расстоянии 20 м от источника излучения. Моделирование аллоксанового диабета осуществляли через месяц после последнего воздействия мШЭИ. 3-я группа – проводилось воздействие мШЭИ, животные были расположены на расстоянии 70 см от источника излучения. Моделирование аллоксанового диабета осуществляли через сутки после последнего воздействия мШЭИ. 4-я группа (плацебо) – проводилось воздействие ШЭИ которое не модулировалось биоструктурой (лазерный луч проходил через пустые предметное и покровное стекла без срезов ПЖ и селезёнки). Животные располагались на расстоянии 70 см от источника излучения. Моделирование аллоксанового диабета в этой группе осуществляли через сутки после последнего воздействия.
В 4-ой группе (плацебо) после превентивного воздействия ШЭИ, не модулированного тканями поджелудочной железы и селезёнки, и последующим введением аллоксана в дозе 200 мг/кг уровень глюкозы в крови на 2-е сутки достоверно отличается от исходного значения (p<0.05). Выживаемость животных составила 80% на 2-е сутки, а к 4-м суткам снизилась до 10%. Это значительно отличалось от показателей выживаемости во 2-ой и 3-ей группах, а так же была ниже показателя выживаемости в контрольной группе (30%) (Рис.1).
Превентивное воздействие мШЭИ значительно влияет на течение экспериментального сахарного диабета у животных во 2-ой и 3-ей группах (Рис.1,2,3, табл.1) и сопровождается защитным, цитопротекторным эффектом (Рис.4). Это наблюдали в обеих указанных группах, несмотря на то, что моделирование экспериментального диабета во 2-ой группе осуществляли, спустя месяц после последнего воздействия мШЭИ, а животные во время воздействия находились на расстоянии 20 м от источника мШЭИ в подвальном помещении лаборатории.
Выживаемость животных 2-ой группы (n=20) составила 90% на 3-е и 4-е сутки с момента введения аллоксана (Рис.1), что значительно отличается от показателей выживаемости в контрольной группе (30%) и 4-ой (плацебо) группе (10%). При этом отмечается достоверное (p<0.05) увеличение уровня глюкозы в крови животных 2-ой группы на 2-е, 3-е и 4-е сутки по сравнению с исходным значением (табл. 1). На 4-е сутки с момента введения аллоксана у 13 животных 2-ой группы (65%) уровень глюкозы в крови был более 14,5 ммоль/л, а у 5 животных (25%) этой группы показатели уровня глюкозы в крови оставались в пределах физиологической нормы (Рис.2). Уровень глюкозы в крови животных 2-ой группы на 4-е сутки достоверно (p<0.05) отличается от исходного значения (табл.1). На 7-е сутки с момента введения аллоксана выживаемость животных во 2-ой группе снизилась до 75% (из 20 животных на фоне выраженной гипергликемии умерло 5 животных) и оставалась на том же уровне до конца наблюдения, в течение 1,5 месяцев (Рис.1). На 8-е сутки с момента введения аллоксана провели эвтаназию 6 животных 2-ой группы, у которых изъяли ткани для патоморфологического исследования. Оставшихся девять крыс наблюдали в течение 1,5 месяцев. Важно отметить, что в период с 8-е по 15-е сутки с момента введения аллоксана у животных с выраженной гипергликемией отмечалось снижение уровня глюкозы в крови (Рис.2). Однако к 18-м суткам с момента введения аллоксана у четверых животных 2-ой группы вновь установилась выраженная стойкая гипергликемия (более 30,6 ммоль/л) которая сохранялась весь период наблюдения. При этом общее состояние этих животных расценивали как удовлетворительное. Аналогичную картину мы наблюдали в наших предыдущих экспериментах [Гаряев, Кокая и др., 2007]. У других оставшихся 5 животных 2-ой группы уровень глюкозы в крови весь период наблюдения оставался в пределах физиологической нормы.
Репродуктивную функцию оценивали у 3 крыс 2-ой группы с выраженной гипергликемией (уровень глюкозы в крови более 30,6 ммоль/л). Крысы дали полноценное многочисленное потомство. Через 1,5 месяца после введения аллоксана у этих животных были изъяты ткани для патоморфологического исследования.
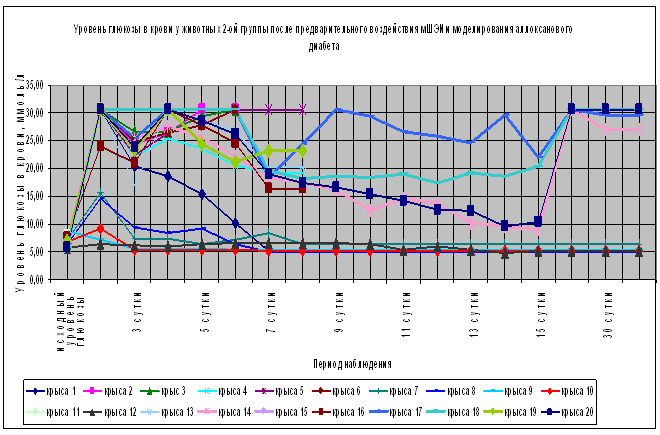
Рис. 2. Влияние превентивного воздействия мШЭИ на течение аллоксанового диабета у крыс 2-ой группы. Животным введен аллоксан в дозе 200 мг/кг спустя месяц после превентивного воздействия мШЭИ. Воздействие проводили в течение 4-х дней по 30 минут. Расстояние от источника излучения 20 м. Животные находились в подвальном помещении. Режим воздействия мШЭИ: 10 минут с использованием ткани поджелудочной железы, 10 минут с использованием ткани селезёнки и 10 минут с использованием ткани поджелудочной железы. Исходный день соответствует дню введения аллоксана.
Более выраженный эффект от превентивного воздействия мШЭИ наблюдали в 3-й группе, которая была расположена на расстоянии 70 см от источника излучения, а моделирование аллоксанового сахарного диабета осуществляли через сутки после последнего воздействия мШЭИ (Рис.1,3, табл.1). В этой группе не было отмечено ни одного случая летального исхода, наблюдалась 100% выживаемость животных весь период наблюдения. У 90% животных уровень глюкозы в крови оставался в пределах физиологической нормы в течение 1,5 месяцев наблюдения, что достоверно (p<0.05) отличается от контрольной, 4-ой и 2-ой групп. Однако у двух крыс из 3-ей группы на 6-е сутки эксперимента был отмечен подъем уровня глюкозы в крови более 20 ммоль/л с последующим снижением до нормальных значений. На 8-е сутки эксперимента провели эвтаназию 6-ти животных 3-ей группы, у которых изъяли ткани для патоморфологического исследования. Репродуктивную функцию оценивали у пяти животных 3-ей группы. Все крысы дали полноценное многочисленное потомство. Через 1,5 месяца после введения аллоксана у шести крыс 3-ей группы были изъяты ткани для патоморфологического исследования. Уровень глюкозы в крови у этих животных был в пределах физиологической нормы. В течение всего периода наблюдения общее состояние всех животных 3-ей группы расценивали как удовлетворительное.
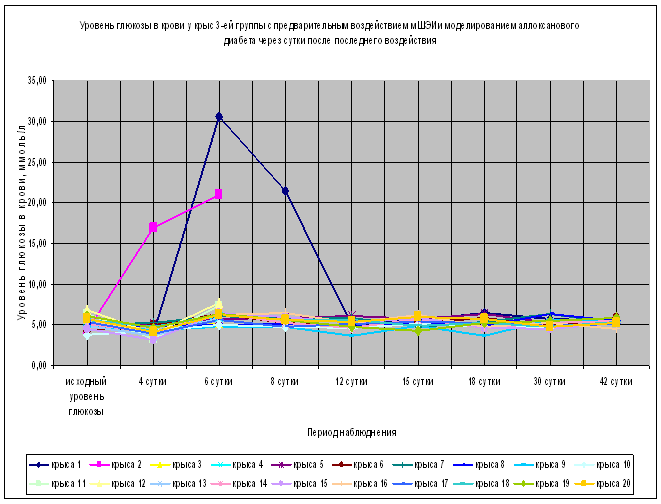
Рис. 3. Влияние превентивного воздействия мШЭИ на течение аллоксанового диабета у крыс 3-ей группы. Животным введен аллоксан в дозе 200 мг/кг спустя сутки после превентивного воздействия мШЭИ. Воздействие проводили в течение 4-х дней по 30 минут ежедневно. Расстояние от источника излучения 70 см. Режим воздействия мШЭИ: 10 минут с использованием ткани поджелудочной железы, 10 минут с использованием ткани селезёнки и 10 минут с использованием ткани поджелудочной железы. Исходный день соответствует дню введения аллоксана.
При гистологическом исследовании препаратов поджелудочной железы в 1-ой, 2-ой и 3-ей группах выявлен ряд особенностей (Рис.4). Гистологическая картина препаратов поджелудочной железы животных контрольной группы характеризовалась выраженными дегенеративными изменениями островков Лангерганса (Рис.4b). Число и размер островков уменьшены, форма их неправильная. Количество β-клеток в островках резко снижено, в большинстве из них отмечалась вакуолизация цитоплазмы, уменьшение размеров ядер, конденсация хроматина, в некоторых клетках кариопикноз. Выявлено наличие лимфоцитарного инфильтрата вокруг и внутри части островков.
Во 2-ой группе на 8-е сутки с момента введения аллоксана гистологическая картина поджелудочной железы характеризовалась деструктивными изменениями разной степени выраженности: островки были уменьшены в размерах, неправильной формы, отмечено снижение количества β-клеток, общая доля инсулярного аппарата в островках резко снижена. Лишь небольшая часть островкового аппарата имела довольно сохранную структуру (Рис. 4с).
Гистологическая картина препаратов поджелудочной железы в 3-ей группе на 8-е сутки с момента введения аллоксана значительно отличалась от контрольной и 2-ой групп. Наряду с патологической картиной изменения островковой ткани поджелудочной железы, в препаратах наблюдали большое количество островков как мелких, так и среднего и крупного размеров с просветлённой цитоплазмой, правильной округлой формой, крупными, округлыми ядрами, содержащими ядрышко (Рис.4е).
Через 1,5 месяца после введения аллоксана гистологическая картина препаратов поджелудочной железы 2-ой группы характеризовалась выраженными дегенеративными изменениями островкового аппарата (Рис.4d).
В отличие от 2-ой и контрольной групп гистологическая картина препаратов в 3-ей группе спустя 1,5 месяца с момента введения аллоксана характеризовалась признаками гипертрофии и гиперплазии поджелудочной железы. Наблюдали большое количество островков разного размера, правильной округлой формы (Рис.4f). Обращало на себя внимание очень большое количество мелких островков и отдельных скоплений β-клеток, а в больших крупных островках наблюдали резкое увеличение числа β-клеток, которые были расположены очень близко друг к другу. Структура островков и отдельных β-клеток была не изменённой, ядра в клетках большие, округлые в которых хорошо определялось ядрышко.
|
|
|
|
|
|
|
|
|
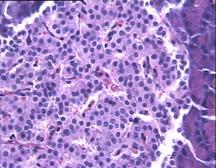
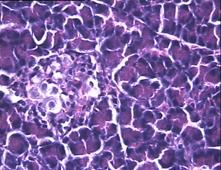
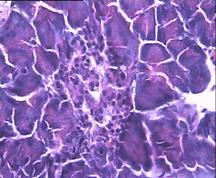
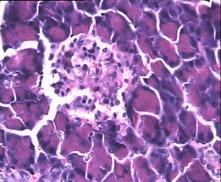
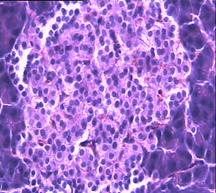
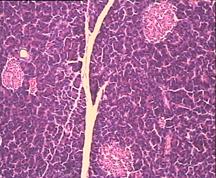
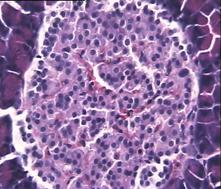
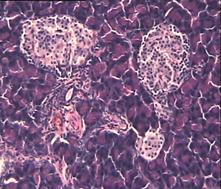
Рис.4. Структура ткани поджелудочной железы, островки Лангерганса: a - интактных крыс; b - 1-ой группы (контроль), после введения аллоксана в дозе 200 мг/кг; c – 2-ой группы на 8-е сутки с момента введения аллоксана в дозе 200 мг/кг. За месяц до моделирования аллоксанового диабета животные этой группы подвергались предварительному воздействию мШЭИ и находились на расстоянии 20 м от источника излучения в подвальном помещении лаборатории; d – 2-ой группы через 1,5 месяца после введения аллоксана в дозе 200 мг/кг. За месяц до моделирования аллоксанового диабета животные этой группы подвергались предварительному воздействию мШЭИ и находились на расстоянии 20 м от источника излучения в подвальном помещении лаборатории; e - 3-ей группы на 8-е сутки с момента введения аллоксана в дозе 200 мг/кг. За сутки до моделирования аллоксанового диабета животные этой группы подверглись предварительному воздействию мШЭИ на расстоянии 70 см от источника излучения; f- 3-ей группы через 1,5 месяца с момента введения аллоксана в дозе 200 мг/кг. Животные этой группы подверглись предварительному воздействию мШЭИ на расстоянии 70 см от источника излучения.
Увеличение 1х400, Увеличение 1х100, окраска гематоксилином и эозином
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Получен положительный эффект от превентивного воздействия мШЭИ на животных 2-ой и 3-ей групп. Различия в динамике уровня глюкозы в крови и выживаемость животных в этих группах указывают на взаимосвязь между сроком воздействия мШЭИ и моделированием аллоксанового сахарного диабета. Биологический защитный эффект дальнодействия при воздействии мШЭИ, обнаруженный в предыдущей работе [Гаряев, Кокая и др., 2007], подтверждается в данном исследовании. Он проявляется в динамике уровня глюкозы в крови у животных 2-ой группы, а также в факте выживаемости животных в этой группе по сравнению с контролем и плацебо. Несмотря на достоверное (p<0.05) увеличение уровня глюкозы в крови у животных 2-ой группы по сравнению с исходным значением и отсутствие достоверных различий между показателями уровня глюкозы в крови у животных 1-ой и 2-ой групп на 2-е, 3-е и 4-е сутки с момента введения аллоксана выживаемость животных в этой группе была высокая. Выраженная гипергликемия у 20% животных данной группы не привела их к гибели, а животные находились в удовлетворительном состоянии весь период наблюдения. Воздействие мШЭИ на животных 3-ей группы способствовало развитию у них устойчивости к действию аллоксану, а уровень глюкозы в крови был в пределах физиологической нормы весь период наблюдения и достоверно (p<0.05) отличался от показателей 1-ой, 2-ой и 4ой групп. Выживаемость в группе составила 100%. Анализируя гистологическую картину препаратов поджелудочной железы в разных группах, можно считать, что превентивное воздействие мШЭИ в 3-ей группе оказало не только цитопротекторный эффект на клетки поджелудочной железы, но и способствовало гипертрофическим и гиперпластическим процессам в ней, которые видимо носили компенсаторный характер. Настоящие экспериментальные результаты находятся в хорошем соответствии с результатами полученными ранее [Гаряев, Кокая и др., 2007].
Таким образом, выявлены три феномена воздействия мШЭИ на крыс с аллоксановым диабетом:
Первый – это фактор выживаемости при выраженной гипергликемии в течение длительного периода наблюдения с сохранением репродуктивной функции у животных.
Второй – в предыдущем исследовании обнаружено, а в настоящем подтверждено, что мШЭИ способствует регенерации поджелудочной железы у больных животных in situ. Третий – предварительное воздействие мШЭИ на животных способствует развитию устойчивости к действию аллоксана.
Зафиксированные эффекты связаны с фундаментальной проблемой «записи» и передачи электромагнитной составляющей генетической информации в ходе постэмбрионального развития с участием волновых процессов в геноме и организме в целом. мШЭИ, параметрически связанное с препарат модулированными фотонами, является, вероятно, носителем и передатчиком информации от биопрепаратов к биосистеме, адресно воспринимающей такую информацию как стратегически управляющую. Вероятно, мШЭИ – это волновой триггер, который запускает «ждущие» регенеративные морфогенетические процессы, информация о которых содержится в геноме каждой клетки. Квантовые механизмы воздействия мШЭИ на эмбриональный и постэмбриональный процессы остаются неизвестными, хотя некоторые идеи в этом направлении высказаны нами ранее [Гаряев, 1994; Гаряев, 1997; Прангишвили, Гаряев и др., 2000(б); Gariaev et al, 2001; Gariaev, 2003] и развиваются в настоящее время. Что касается защитного и цитопротекторного действия мШЭИ, то эта область исследований имеет огромные перспективы.
Возможно, определенную роль в обнаруженных защитно-регенерационных проявлениях играет фактор так называемого «слабого воздействия» [Чукова, 2002]. В этом плане можно предполагать, что обнаруженные нами эффекты носят эндоэргический характер, когда даже слабо поглощаемая препаратами энергия когерентного поляризованного лазерного излучения идет на увеличение свободной энергии Гельмгольца, аккумулированной в химических связях метаболитов препаратов поджелудочной железы и селезенки. Например, атомы информационных макромолекул (ДНК, РНК, белки), поглощая свет, вместе с энергией квантов света приобретают и их момент количества движения, что создает инверсную заселенность ядерных зеемановских уровней. Происходит так называемая химическая поляризация ядер. Таким образом, биохимические реакции в препаратах, запущенные поляризованным лазерным излучением, могут генерировать электромагнитные радиочастотные колебания. В этой ситуации препараты поджелудочной железы и селезенки выступают в роли своеобразной молекулярной радиостанции, где каждый вид молекул имеет свои характерные частоты, которые, ввиду наличия в эксперименте широкополосного радиоизлучения газового разряда лазера, могут усиливаться благодаря стохастическому резонансу.
На основании полученных экспериментальных данных предлагается попытаться разработать технологию позволяющую выработать устойчивость животных к токсическому действию гептила в эксперименте. Мы предполагаем, что устойчивость к гептилу и многим другим токсическим веществам можно выработать, воздействуя на стратегические метаболические вектора, важнейшие из которых – функции генетического аппарата на квантовом уровне http://www.wavegenetics.jino-net.ru/.
Общее обсуждение экспериментальных результатов. Теоретические модели.
Полученные данные по значимости шире, чем демонстрация возможности волнового защитного антидот эффекта и требует теоретического осмысления, поскольку затрагивают стратегические (квантовые) механизмы работы генетического аппарата многоклеточных биосистем. Мы предлагаем три формализованные гипотезы волновых событий при считывании с биоструктуры-донора генетико-матаболической волновой информации, дистантной адресной передаче ее, введении в биосистему-акцептор и управлению ее метаболизмом.
1.Эндогенные поляризационно-голографические события в биосистемах.
Волновые информационные сценарии, разворачивающиеся как в самой биосистеме, так и в процессе считывания их лазерным лучом, на первом этапа происходят на фотонном уровне. Этот уровень рассмотрим подробно. В наших работах [Прангишвили, Гаряев и др., 2000(а); Прангишвили, Гаряев и др., 2000(б)] были даны двух- и трехмерные модели биоголографического управления построением пространственной структуры многоклеточных организмов в процессе эмбриогенеза. В первом приближении при условии относительно стационарных состояний в биосистемах (финальные стадии морфогенезов), эти модели достаточно реалистичны. Однако в живых организмах статика и динамика парадоксально совмещены. Взрослый организм пространственно относительно статичен в макро масштабе и существенно меняется в этом плане только на стадиях глубокого старения. Вместе с тем, эта статика обеспечивается внутренней пространственно-временной динамикой метаболических процессов на микроуровне организации биосистем. Это обусловлено тем, что процесс метаболизма является мобильной совокупностью биохимико-биофизических пространственно-временных преобразований микроструктуры организма. С учетом нестационарности структуры биосистем предлагается более развитая модель эндогенных информационных поляризационно-голографических управляющих процессов в многоклеточных организмах, реализуемых прежде всего на уровне генома. Модель отображает биоголографический аспект метаболизма в целом и поэтому включает в себя биоморфогенез в качестве его частного случая. Модель использует существующий физико-математический формализм для поляризационной голографии, но экстраполирует его на вероятные эндогенные аналогичные процессы в генетическом аппарате многоклеточных организмов.
В
основу модели заложены также наши экспериментальные исследования с
использованием специального двух поляризационного He-Ne лазера (![]() = 632,8нм), имеющего
две ортогональные, связанные между собой, оптические моды, о чем говорилось
выше. При взаимодействии лазерного пучка такого квантового генератора с
веществом, в режиме динамического голографирования на встречных пучках,
происходят акты одновременной записи-считывания неизвестной ранее информации о
динамических вращательно-колебательных процессах на оптическом и
атомно-молекулярном уровнях. Особенно интересны полученные таким путем сведения
о генетических структурах и/или о живых клетках. Все информационные структуры
организмов, включая ДНК, РНК и белки, обладают оптической активностью, то есть
способностью вращать плоскость поляризации света и дихроичностью – разностью
поглощений право- и лево поляризованного света. Модуляции поляризации,
коррелирующие со структурно-функциональным состоянием того или иного
метаболита, выступают как уникальное по своей емкости вместилище информации о
метаболизме и его динамике. И вместе с тем – это канал межклеточных фотонных
биознаковых контактов. Такие особенности процессов, в
поляризационно-голографическом варианте, по-видимому, присущи работе генома как
биокомпьютера. Это позволяет моделировать их с использованием упомянутого
лазера. Он способен к поляризационно-голографической записи, считыванию,
дистантной передаче и введению волновой командной генетико-метаболической
информации от одной биосистемы к другой. Кроме того, такой лазер осуществляет
конверсию зондирующих биосистему фотонов в широкополосный электромагнитный
спектр с частотами от 2
= 632,8нм), имеющего
две ортогональные, связанные между собой, оптические моды, о чем говорилось
выше. При взаимодействии лазерного пучка такого квантового генератора с
веществом, в режиме динамического голографирования на встречных пучках,
происходят акты одновременной записи-считывания неизвестной ранее информации о
динамических вращательно-колебательных процессах на оптическом и
атомно-молекулярном уровнях. Особенно интересны полученные таким путем сведения
о генетических структурах и/или о живых клетках. Все информационные структуры
организмов, включая ДНК, РНК и белки, обладают оптической активностью, то есть
способностью вращать плоскость поляризации света и дихроичностью – разностью
поглощений право- и лево поляризованного света. Модуляции поляризации,
коррелирующие со структурно-функциональным состоянием того или иного
метаболита, выступают как уникальное по своей емкости вместилище информации о
метаболизме и его динамике. И вместе с тем – это канал межклеточных фотонных
биознаковых контактов. Такие особенности процессов, в
поляризационно-голографическом варианте, по-видимому, присущи работе генома как
биокомпьютера. Это позволяет моделировать их с использованием упомянутого
лазера. Он способен к поляризационно-голографической записи, считыванию,
дистантной передаче и введению волновой командной генетико-метаболической
информации от одной биосистемы к другой. Кроме того, такой лазер осуществляет
конверсию зондирующих биосистему фотонов в широкополосный электромагнитный
спектр с частотами от 2![]() до 0 по механизмам
локализации-делокализации фотонов. При этом, видимо, сохраняется квантово
нелокальная (телепортационная) поляризационная связь по всему набору частот,
включая радиоволновые. Использование такого лазера как считывающе-передающую
фотонно-радиоволновую систему, имитирующую аналогичные волновые
биокомпьютерные знаковые нелокальные процессы межклеточных коммуникаций, дало
возможность осуществить дальнюю волновую передачу управляющей
генетико-метаболической информации от биосистем-доноров к
биосистемам-акцепторам. В свете этого факта представляется существенным
попытаться дать версию более развитого формализма биознаковых
фотоннно-поляризационно-голографических процессов в хромосомном аппарате высших
биосистем, тем более, что радиоволновой эквивалент этих процессов обладает ярко
выраженными морфогенетическими потенциями.
до 0 по механизмам
локализации-делокализации фотонов. При этом, видимо, сохраняется квантово
нелокальная (телепортационная) поляризационная связь по всему набору частот,
включая радиоволновые. Использование такого лазера как считывающе-передающую
фотонно-радиоволновую систему, имитирующую аналогичные волновые
биокомпьютерные знаковые нелокальные процессы межклеточных коммуникаций, дало
возможность осуществить дальнюю волновую передачу управляющей
генетико-метаболической информации от биосистем-доноров к
биосистемам-акцепторам. В свете этого факта представляется существенным
попытаться дать версию более развитого формализма биознаковых
фотоннно-поляризационно-голографических процессов в хромосомном аппарате высших
биосистем, тем более, что радиоволновой эквивалент этих процессов обладает ярко
выраженными морфогенетическими потенциями.
Запишем векторный дифракционный интеграл Кирхгофа в параксиальном приближении, описывающий волновое, например фотонное поле, сформированное нестационарным фрагментом биоструктуры. Такое фотонное поле может излучаться жидкокристаллическим континуумом хромосом (ЖКХ) in vivo. Вид такого излучения может выражаться следующим соотношением:
E![]() (x,y,z,
(x,y,z,![]() ,t)
,t)![]()
![]()
![]() , (1)
, (1)
где c - скорость света; ![]() частота; x
частота; x![]()
![]() и x, y, z, t – соответственно пространственно-временные координаты
точки фрагмента ЖКХ и точки наблюдения; r –
расстояние между этими точками; S
и x, y, z, t – соответственно пространственно-временные координаты
точки фрагмента ЖКХ и точки наблюдения; r –
расстояние между этими точками; S![]() - пространственно-временной
интервал, занимаемый ЖКХ; dS=dx
- пространственно-временной
интервал, занимаемый ЖКХ; dS=dx![]()
В
уравнении (1) E![]() - распределение амплитуды
поля за ЖКХ. Если рассматривать процессы голографирования образцов
биотканей-доноров используемым лазером, то это поле имеет место для каждой
поляризационной моды, которые между собой ортогональны и независимы до тех
пор, пока не произойдет поворот плоскостей их первоначальных положений
векторов поляризованных одночастотных и несколько смещенных по частоте друг
относительно друга волн со средней частотой
- распределение амплитуды
поля за ЖКХ. Если рассматривать процессы голографирования образцов
биотканей-доноров используемым лазером, то это поле имеет место для каждой
поляризационной моды, которые между собой ортогональны и независимы до тех
пор, пока не произойдет поворот плоскостей их первоначальных положений
векторов поляризованных одночастотных и несколько смещенных по частоте друг
относительно друга волн со средней частотой ![]() , распространяющихся вдоль
оси z, с вектором Джонса. Еще раз напомним, что для
хромосом характерна высокая оптическая активность, выражающаяся в дисперсии
оптического вращения и круговом дихроизме, что является необходимым условием
применения формализма.
, распространяющихся вдоль
оси z, с вектором Джонса. Еще раз напомним, что для
хромосом характерна высокая оптическая активность, выражающаяся в дисперсии
оптического вращения и круговом дихроизме, что является необходимым условием
применения формализма.
E![]() =E
=E![]() ,
0
,
0![]() (2)
(2)
Поле E![]() проходит через
нестационарный фрагмент ЖКХ с матрицей Джонса
проходит через
нестационарный фрагмент ЖКХ с матрицей Джонса
M![]() (x
(x![]() ,y
,y![]() ,z
,z![]() ,t
,t![]() )=
)=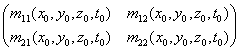
Для упрощения будем считать, что нестационарность ЖКХ не является функцией частоты просвечивающего света.
Обе поляризационные моды когерентного света лазера деполяризуются гено-знаковой акустикой ЖКХ и частично эллиптически поляризуются. При этом они могут интерферировать с образованием спекл-структуры, а суммарная их интенсивность «перетекает» из моды в моду по пути, постулированному ранее [Прангишвили, Гаряев и др., 2000(а)]. Это, в свою очередь, приводит к модуляции радиоволн, образующихся из хромосомных фотонов по механизму их делокализации [Прангишвили, Гаряев и др., 2000(б)].
Модифицированный вектор Джонса каждой из прошедших ортогонально поляризованных волн непосредственно за объектом может быть представлен в виде частично когерентных ортогональных компонент эллиптической поляризации
E![]() (3)
(3)
где ![]() знак
некогерентного суммирования амплитуд, который введен для частично
поляризованного света;
знак
некогерентного суммирования амплитуд, который введен для частично
поляризованного света; ![]() комплексная амплитуда компоненты одного
базиса;
комплексная амплитуда компоненты одного
базиса; ![]() комплексная
амплитуда компоненты другого, ортогонального ему и некогерентного.
комплексная
амплитуда компоненты другого, ортогонального ему и некогерентного.
В
биологической системе в составе ЖКХ (при наличии только одной поляризационной
компоненты) используем в качестве гипотетической опорной волну, прошедшую,
например, через бесконечно узкий временной затвор, имеющий ![]() - образную
характеристику временного пропускания. Такой затвор полностью деполяризует
изначально поляризованную волну. Полученная позади затвора волна обладает
сплошным спектром во всем диапазоне с равномерно распределенной спектральной
плотностью, а модифицированный вектор опорной волны имеет вид ортогонального
базиса эллиптической поляризации:
- образную
характеристику временного пропускания. Такой затвор полностью деполяризует
изначально поляризованную волну. Полученная позади затвора волна обладает
сплошным спектром во всем диапазоне с равномерно распределенной спектральной
плотностью, а модифицированный вектор опорной волны имеет вид ортогонального
базиса эллиптической поляризации:
Eоп = ![]() ,
(4)
,
(4)
где ![]() амплитуды;
амплитуды;
![]() соответственно
начальные фазы двух взаимно некогерентных компонент.
соответственно
начальные фазы двух взаимно некогерентных компонент.
Для нашего случая, где используется сразу две поляризационные компоненты, не требуется вышеуказанного допущения о наличии бесконечно узкого временного затвора и тогда суммарное поле в плоскости поляризационной голограммы имеет вид:
E![]() E
E![]() + E
+ E![]() (5)
(5)
E![]()
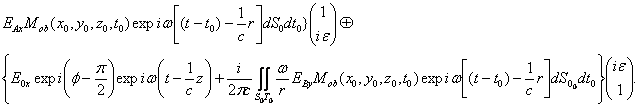
Реальная часть последнего уравнения (5) представляет напряженность электрического вектора суммарной волны.
Re (E![]() ) = p cos
) = p cos![]() t + g sin
t + g sin![]() t,
(6)
t,
(6)
Параметры суммарного эллипса p и g определяются через компоненты эллипса поляризации каждого из базисов А и В, как в работе [14]
p = Re(E![]() )
)![]()
![]() Re(E
Re(E![]() )
)![]() = p
= p![]()
![]() p
p![]()
g = Im(E![]() )
)![]()
![]() Im (E
Im (E![]() )
)![]() = g
= g![]()
![]() g
g![]() (7)
(7)
Эндогенная биологическая регистрация поля суммарной волны (5) в составе ЖКХ, как базовом элементе ДНК-волнового биокомпьютера, предполагает наличие в организмах поляризационно-чувствительной среды, которая, так же как и нестационарный фрагмент биологического объекта, например ЖКХ, спектрально не селективен во всем диапазоне действующих частот.
За счет
поляризационных характеристик индуцирующего света в светочувствительной
регистрирующей среде ЖКХ наводятся фотоанизотропия и фотогиротропия. Для
описания векторного фотоотклика поляризационно-чувствительной среды введены
функции изотропной ![]() , анизотропной
, анизотропной ![]() - и гиротропной
- и гиротропной ![]() -
реакций, которые постоянны для всех частот действующего излучения. Используя
матрицы Джонса и правила их построения для случая частично поляризованного
индуцирующего излучения, для результирующей матрицы Джонса получаем
-
реакций, которые постоянны для всех частот действующего излучения. Используя
матрицы Джонса и правила их построения для случая частично поляризованного
индуцирующего излучения, для результирующей матрицы Джонса получаем
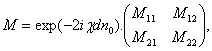 (8)
(8)
где
![]()
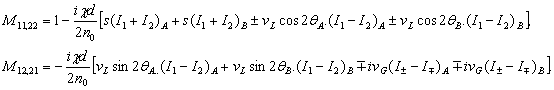
![]() В (8)
В (8) ![]() длина
исходной просвечивающей эндогенной волны (например, фотонное излучение хромосом
in vivo);
длина
исходной просвечивающей эндогенной волны (например, фотонное излучение хромосом
in vivo);
![]() -
толщина регистрирующей ЖКХ;
-
толщина регистрирующей ЖКХ; ![]() - комплексный коэффициент преломления
ЖКХ в исходном, необлученном состоянии;
- комплексный коэффициент преломления
ЖКХ в исходном, необлученном состоянии; ![]() и
и ![]() - первый параметр
Стокса,
- первый параметр
Стокса,
![]() и
и ![]() - второй
параметр Стокса,
- второй
параметр Стокса, ![]() и
и ![]() - четвертый параметр
Стокса для
- четвертый параметр
Стокса для ![]() и
и ![]() компонент;
компонент; ![]() и
и ![]() - углы
ориентации большой оси эллипса поляризации соответственно для
- углы
ориентации большой оси эллипса поляризации соответственно для ![]() - и
- и ![]() -
компонент, отсчитываемые против часовой стрелки относительно оси
-
компонент, отсчитываемые против часовой стрелки относительно оси ![]() .
.
Выразив
в (8) параметры Стокса через параметры p![]() p
p![]() g
g![]() ,g
,g![]() для матрицы голограмм,
представленной в виде суммы трех матриц во всем диапазоне действующих
частот, получим
для матрицы голограмм,
представленной в виде суммы трех матриц во всем диапазоне действующих
частот, получим
![]() (9)
(9)
где ![]() -
матрица, описывающая недифрагированный пучок,
-
матрица, описывающая недифрагированный пучок,
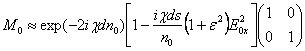 ;
(10)
;
(10)
![]() -
матрица, описывающая мнимое изображение,
-
матрица, описывающая мнимое изображение,
![]()
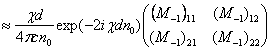 (11)
(11)
с матричными элементами
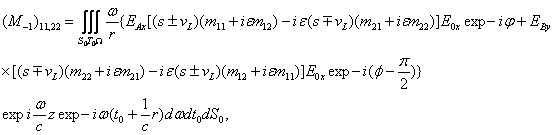
![]()
![]()
![]()
![]()
![]() -
матрица описывающая действительное изображение
-
матрица описывающая действительное изображение
![]()
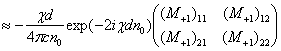 (12)
(12)
с матричными элементами
![]()
![]()
![]()
![]()
![]()
![]()
Здесь
![]() -
зависящие от координат и времени элементы двумерной матрицы нестационарного
фрагмента ЖКХ. Исходя из условия отбора оптимальных гомеостатических
состояний в биологических объектах, соотношения между функциями реакции среды
могут быть следующими:
-
зависящие от координат и времени элементы двумерной матрицы нестационарного
фрагмента ЖКХ. Исходя из условия отбора оптимальных гомеостатических
состояний в биологических объектах, соотношения между функциями реакции среды
могут быть следующими:
![]()
![]() (13)
(13)
и выражения (11) и (12) значительно упрощаются. Условия (13) выполняются с большой точностью для очень большого класса поляризационно чувствительных сред.
При
выполнении условий (13) матрицы ![]() и
и ![]() имеют следующий вид:
имеют следующий вид:
![]() (14)
(14)
![]() (15)
(15)
В
(14) и (15) выделена матрица ЖКХ ![]() , а через
, а через ![]() обозначена
следующая матрица
обозначена
следующая матрица
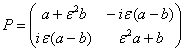 ,
,
где
![]()
![]() ;
;
![]() -
эрмитово сопряженные матрицы.
-
эрмитово сопряженные матрицы.
При условии эндогенного освещения полученной голограммы реконструирующей эндогенной или экзогенной по отношению к биосистеме неполяризованной волной с комплексными амплитудами
![]() и
частотой
и
частотой ![]()
![]() (16)
(16)
прошедшая через биологическую голограмму волна формируется в виде
![]() (17)
(17)
где ![]() - размер
фрагмента голограммы ЖКХ;
- размер
фрагмента голограммы ЖКХ; ![]() - расстояние между точкой на
поверхности голограммы и точкой наблюдения.
- расстояние между точкой на
поверхности голограммы и точкой наблюдения.
Затем,
последовательно подставляя в (17) выражения для матриц (10), (14) и (15),
определим сформированные голограммой нулевое, мнимое и действительное
изображения. И только теперь определим, какую эндогенную и/или экзогенную
волну для организма необходимо использовать в качестве реконструирующей, чтобы
получить в мнимом виде восстановление нужного нам фрагмента волнового образа
формирующейся биосистемы. Для этого необходимо определить собственные векторы и
соответствующие им собственные значения матрицы ![]() . Оказывается, что с
точностью до постоянного множителя собственные векторы матрицы
. Оказывается, что с
точностью до постоянного множителя собственные векторы матрицы ![]() суть
суть ![]() и
и ![]() с
соответствующими собственными
значениями
с
соответствующими собственными
значениями
![]() и
и
![]() .
.
Отсюда следует, что восстановление производится волной, идентичной использованной, при записи опорной волной. А так как, вероятно, в биосистемах на уровне ЖКХ запись и восстановление происходят или одновременно, или с соблюдением последнего условия, то реконструированное мнимое изображение соответствует истинному, и оно не подвержено никаким искажениям. Последнее принципиально важно для сохранения волновых образов-векторов морфогенеза, компенсирующих физиолого-биохимическую и механическую нестационарность биосистемы в целом и ее ЖКХ, в частности. Тем не менее, нестабильность стратегических фотонных образов структуры организма будет иметь место, но на больших временных отрезках при старении организма и его патологических состояниях, например, в случае канцерогенеза.
Для прошедшей
без дифракции волны нулевое изображение имеет вид:![]()
(18),
а мнимое и действительное изображения соответственно представляются в виде
![]()
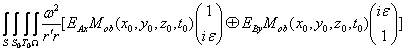
![]()
![]() (19)
(19)
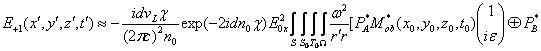
![]() (20)
(20)
где
![]() ,
,
![]()
Интегралы,
входящие в (19) и (20), в работе [21] решены в линейном приближении для
расстояний ![]() и
и ![]() и для бесконечно больших
областей интегрирования
и для бесконечно больших
областей интегрирования ![]() . Интегралы по
. Интегралы по ![]() и
и ![]() имеют
характер соответственно пространственной и временной
имеют
характер соответственно пространственной и временной ![]() -функции. Окончательные
выражения, приводят к следующим выражениям для сформированных
пространственно-временной поляризационной голограммы. Для сформированного
мнимого изображения при
-функции. Окончательные
выражения, приводят к следующим выражениям для сформированных
пространственно-временной поляризационной голограммы. Для сформированного
мнимого изображения при ![]() из (19) имеем
из (19) имеем
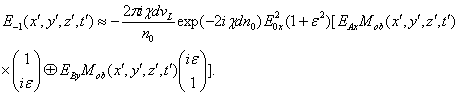 (21)
(21)
Анализ последнего соотношения показывает, что с точностью до множителя в нем отображено полное восстановление как пространственно-временной структуры, так и поляризационных характеристик поля его нестационарной объектной волны, прошедшей, например, через ЖКХ. Эти фотонные и/или радиоволновые динамичные структуры, вероятно, используются многоклеточными организмами для собственной организации в собственных пространстве-времени, поскольку эти структуры-образы полностью сохраняет истинный калибровочный масштаб без искажений, накладываемых нестационарностью биосистем, и воспроизводят их его в адекватных для развивающегося и взрослого организма измерениях. По реконструированным волновым градиентам считанных поляризационных голограмм осуществляется 4-мерная организация метаболических потоков, клеточной архитектоники и морфогенетических движений в ходе эмбриогенеза, а также частичная регенерации биосистем при их повреждениях. Иными словами, происходит калибровка динамичного потенциального пространства-времени биосистемы.
Из (20) для действительного изображения при ![]() имеем
имеем
![]()
![]() .
(22)
.
(22)
Из (22)
следует, что на расстоянии ![]() , симметрично мнимому изображению
(19) относительно голограммы, формируется изображение с псевдоскопической
пространственной структурой объектного фрагмента поля ЖКХ. При этом происходит
обращение его временного профиля с временной задержкой, вызванной прохождением
светом расстояния
, симметрично мнимому изображению
(19) относительно голограммы, формируется изображение с псевдоскопической
пространственной структурой объектного фрагмента поля ЖКХ. При этом происходит
обращение его временного профиля с временной задержкой, вызванной прохождением
светом расстояния ![]() , равного расстоянию от точки наблюдения
до действительного изображения, с преобразованием состояния поляризации,
определяемым видом матриц
, равного расстоянию от точки наблюдения
до действительного изображения, с преобразованием состояния поляризации,
определяемым видом матриц ![]() и
и ![]() .
.
Поляризационно-голографическое биоуправление мы понимаем как эндогенное или искусственное изменение состояния и структуры клеток реципиента в результате управляющих голографических воздействий со стороны донора. В нашем случае голографический сигнал, модулированный здоровыми клетками донора, транслируется и записывается на больных клетках реципиента в виде голограммы. Далее процесс управления происходит следующим образом. Вначале с измененных клеток реципиента под действием восстанавливающей волны считывается голографическое изображение здоровых клеток донора. Оно реконструируется в терагерцовом диапазоне волн в виде объемного изображения, охватывающего каждую клетку реципиента вместе с ее наполнением.
В принципе существуют два вида источников восстанавливающей волны. Первый из них – эндогенный. В этом случае процессы протекают за счет внутренних резервов, т.е. «внутреннего» излучения соседних клеток. Второй вид – экзогенная реконструкция, когда источниками являются внешние излучатели. Оба вида источников «работают» в клетках реципиента и действуют одновременно и постоянно, восстанавливая и дополняя одно и то же изображение здоровых клеток донора.
По градиентам интенсивности восстановленного изображения донорной клетки, как по чертежу, происходит рост и регенерация больных клеток реципиента. Клетки реципиента играют при этом роль фотопластинки, на которой записана голограмма здоровых клеток донора. Процессы роста и регенерации, протекающие аналогично процессам фототропизма, занимают определенное время. В результате «больные» клетки реципиента частично переходят в разряд здоровых, а частично разрушаются. Продукты распада разрушенных клеток выводятся из организма реципиента.
Таким образом, в процессе замещения клеток реципиента здоровыми клетками, аналогичными образу донора, происходит поляризационно-голографическое управление, состоящее в «прорастании» биомассы больных клеток реципиента в предложенную им динамическую голографическую форму клеток здорового донора. Следовательно, в процессе управления форма и динамическое состояние клеток реципиента постепенно меняются под действием управляющего сигнала – «установки», полученной от донора.
Разумеется, такая «установка» намного сложнее той «установки», которая встречается в простых системах управления. Она задает пространственные распределения терагерцового сигнала, по градиентам которого происходит рост и формирование живых клеток в реципиенте. Таким образом, процессы роста и формообразования реципиентных клеток протекают по биохимическим законам, управляющим их жизнедеятельностью, а сигнал «установки» только задает программу роста структуры молодых здоровых клеток и модуляцию протекающих в них процессов.
Для более полного описания механизма внутренней работы голографического циркулярно-поляризованного информационно-лазерного преобразования в живом организме необходимо понимание основ поляризационно-динамической теории голографии и обмена информацией между живой здоровой клеткой донора и всеми другими клетками, входящими в состав органов и тканей больного организма. При таком описании мы используем работу классика в области динамической голографии Ю.Н. Денисюка [Денисюк, 1974], разработавшего основы регистрации и реконструкции голографического построения изображений материальных структур. Нам удалось экспериментально доказать, что помимо регистрации и реконструкции изображений возможно и внутриклеточное видоизменение биологических структур реципиента по схеме восстановленного изображения донора, записанного на голографических структурах реципиента.
Трансляция модуляционной информации от донора к реципиенту происходит посредством прямолинейно распространяющихся продольных взаимно проникающих волн, несущих многоуровневую модуляционную информацию. Для ближней зоны передачи информации можно применять понятие клеточных ядер - оптических поляроидов, а для дальней – понятие квази объектива (см. предыдущую статью данного сборника). Рассмотрим описание такого процесса, предложенное для регистрации цветных голограмм без использования лазеров [Александров, 1998]. Адаптируя его к биологической системе, опишем необходимые условия для реализации некогерентного поляризационно-голографического метода управления. Следует заметить, что в этих системах при микроскопическом их изучении, давно обнаружили как микроскопические поляризаторы, тое есть ядра клеток, так и оптически активные белковые вещества, которые вращают плоскость поляризации проходящих через них излучений. Эти процессы давно известны исследователям [Bischof, 1995], однако до сих пор наблюдаемое явление не было ни объяснено, ни использовано.
Пусть под углом ![]() от произвольной точки биологического
объекта-донора свет попадает на первый внутриклеточный поляризатор и далее
проходит через оптически активную среду, например, через биологический белок.
Пройдя через второй клеточный поляроид, свет падает на регистрирующую биологическую
среду реципиента.
от произвольной точки биологического
объекта-донора свет попадает на первый внутриклеточный поляризатор и далее
проходит через оптически активную среду, например, через биологический белок.
Пройдя через второй клеточный поляроид, свет падает на регистрирующую биологическую
среду реципиента.
После прохождения первого поляроида амплитуду волны можно записать в
виде ![]() , где
, где ![]() - фаза волны;
- фаза волны; ![]() -
вещественная амплитуда световой волны после прохождения поляризатора
-
вещественная амплитуда световой волны после прохождения поляризатора ![]() - угол,
определяющий положение первого поляризатора в выбранной системе координат.
Далее свет проходит оптически активную среду (белок) и плоскость его
поляризации поворачивается на угол
- угол,
определяющий положение первого поляризатора в выбранной системе координат.
Далее свет проходит оптически активную среду (белок) и плоскость его
поляризации поворачивается на угол ![]() , который зависит от угла преломления
, который зависит от угла преломления ![]() при
фиксированной толщине
при
фиксированной толщине ![]() оптически активного белка:
оптически активного белка:
![]() (3)
(3)
где ![]() -
постоянная вращения вектора поляризации. Из этой формулы видно, что поворот
векторов поляризации не зависит от длины зондирующей волны (в формулу не входит
длина волны
-
постоянная вращения вектора поляризации. Из этой формулы видно, что поворот
векторов поляризации не зависит от длины зондирующей волны (в формулу не входит
длина волны ![]() ), а зависит только от вращательной
степени активности среды
), а зависит только от вращательной
степени активности среды ![]() , ее толщины
, ее толщины ![]() и угла преломления
этой среды
и угла преломления
этой среды ![]() .
.
С
учетом введенных обозначений амплитуда световой волны имеет вид: ![]() где
где ![]() -
передаточный коэффициент.
-
передаточный коэффициент.
После
прохождения через второй биологический поляризатор скалярная амплитуда волны в
плоскости поляризации имеет вид ![]()
где ![]() передаточный
коэффициент,
передаточный
коэффициент, ![]() - угол, определяющий положение второго
поляроида в выбранной системе координат.
- угол, определяющий положение второго
поляроида в выбранной системе координат.
Тогда интенсивность света будет равна
![]() ,
(4)
,
(4)
где ![]() -
передаточный коэффициент для интенсивности,
-
передаточный коэффициент для интенсивности, ![]() - интенсивность света, рассеянного
некоторой точкой, расположенной на зондируемом биологическом объекте - доноре.
Подставив выражение (3) в (4), получим
- интенсивность света, рассеянного
некоторой точкой, расположенной на зондируемом биологическом объекте - доноре.
Подставив выражение (3) в (4), получим
![]() .
(5)
.
(5)
Анализируя полученное уравнение (5), можно заметить, что закон распределения
интенсивности поляризационных полос в этой формуле аналогичен известному закону
распределения интенсивности интерференционных полос в зонной решетке Габора,
т.е. представляет собой осевую голограмму точки объекта. Кроме того, величина
интенсивности зависит от величины угла ![]() , а при
, а при ![]() = const,
и
= const,
и ![]() само
распределение интенсивности света должно иметь вид чередующихся темных и
светлых поляризационных колец с переменным периодом.
само
распределение интенсивности света должно иметь вид чередующихся темных и
светлых поляризационных колец с переменным периодом.
Если учесть, что
![]() зависит
от величины коэффициента преломления оптически активной среды, в качестве
которой в нашем случае выступает белок клетки, то можно записать значение
зависит
от величины коэффициента преломления оптически активной среды, в качестве
которой в нашем случае выступает белок клетки, то можно записать значение ![]() :
:
![]() .
(6)
.
(6)
С учетом выражения (4) перепишем уравнение (5) в виде
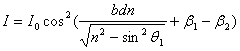 .
.
Пренебрегая
рефракцией, то есть при ![]() , получим выражение для
, получим выражение для ![]() :
:
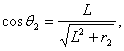 (7)
(7)
где ![]() -
расстояние от точки биологического объекта (донора) до плоскости регистрации
голограммы (реципиента), а
-
расстояние от точки биологического объекта (донора) до плоскости регистрации
голограммы (реципиента), а ![]() - расстояние от осевой линии,
проходящей через центр регистратора голограммы до точки, в которую попадает
луч, исходящий от точки биологического объекта-донора.
- расстояние от осевой линии,
проходящей через центр регистратора голограммы до точки, в которую попадает
луч, исходящий от точки биологического объекта-донора.
Предположим, что поляризационные элементы установлены параллельно, то есть
![]() . Тогда,
подставив (6) в (4), получим
. Тогда,
подставив (6) в (4), получим
![]() .
(8)
.
(8)
Из последнего выражения видно, что закон распределения интенсивности света в плоскости регистрации реципиента есть функция, зависящая от положения голографируемой точки, находящейся внутри или на поверхности биологического объекта-реципиента.
При нормальном падении луча света на первый поляроид (![]() ), когда
наблюдается максимальная величина интенсивности, угол поворота второго
поляроида
), когда
наблюдается максимальная величина интенсивности, угол поворота второго
поляроида ![]() должен удовлетворять соотношению
должен удовлетворять соотношению ![]() . Здесь
. Здесь ![]() - угол
поворота поляризации света при прохождении нормально падающего луча через
оптически активный белок клетки.
- угол
поворота поляризации света при прохождении нормально падающего луча через
оптически активный белок клетки. ![]() - число поворотов плоскости
поляризации на
- число поворотов плоскости
поляризации на ![]() при прохождении нормально падающего
луча через оптически активный белок клетки-реципиента. При этом распределение
интенсивности света исходящего от донора в плоскости регистрации голограммы,
где находится реципиент, примет вид
при прохождении нормально падающего
луча через оптически активный белок клетки-реципиента. При этом распределение
интенсивности света исходящего от донора в плоскости регистрации голограммы,
где находится реципиент, примет вид ![]() откуда, используя формулы (4) и (7),
получим
откуда, используя формулы (4) и (7),
получим
![]() (9)
(9)
Внутри окружности радиуса ![]() расположено
расположено ![]() светлых (или
темных) колец, определяемых по формуле
светлых (или
темных) колец, определяемых по формуле ![]() . Отсюда радиус
. Отсюда радиус ![]() го кольца можно
определить по формуле
го кольца можно
определить по формуле ![]() .
.
Для сравнения приведем распределение интенсивности в обычной когерентной осевой голограмме точки, получаемой в результате интерференции объектной сферической и опорной плоской волн
![]() (10)
(10)
где ![]() Сравнивая
выражения (10) и (9), можно отметить, что аргументы косинусов отличаются
масштабным множителем, пропорциональным
Сравнивая
выражения (10) и (9), можно отметить, что аргументы косинусов отличаются
масштабным множителем, пропорциональным ![]() характеризующим вклад оптически
активной среды в общий путь света.
характеризующим вклад оптически
активной среды в общий путь света.
В декартовой системе координат распределение интенсивности для голограммы точки (8) можно записать в виде
![]() (11)
(11)
где ![]()
Голограммой места донора является суперпозиция распределений (11), а распределение интенсивности в голограмме реципиента имеет вид
![]() (12)
(12)
Отсюда можно записать импульсную характеристику (или голографическую функцию размытия точки) следующим образом:
![]() .
(13)
.
(13)
Голографическую передаточную функцию можно определить, исходя из Фурье-преобразования выражения (13). Напомним, что в эту формулу не входит длина волны зондирующего сигнала, так что эта волна может быть выбрана в широком спектре волн светового, электромагнитного и акустического диапазонов. Полученная голограмма содержит полную информацию о пространственных координатных характеристиках голографируемого объекта-донора или о пространственном распределении всех точек донора относительно плоскостей регистрации голограмм реципиента.
Таким образом, полученное решение задачи, вообще говоря, аналогично традиционному. В то же время предложенный метод принципиально отличается от известных интерференционных методов и имеет определенные преимущества.
Во-первых, вместо некоторой длины волны ![]() с ее монохроматичностью и
когерентностью используется дисперсионная вращательная способность оптически
активной среды
с ее монохроматичностью и
когерентностью используется дисперсионная вращательная способность оптически
активной среды ![]() и пространственная
локально-распределенная поляризационная фильтрация. Этого вполне достаточно,
чтобы при наличии собственных движений клетки-донора в условиях некогерентного
широкоспектрального облучения реципиента была записана
поляризационно-динамическая голограмма донора.
и пространственная
локально-распределенная поляризационная фильтрация. Этого вполне достаточно,
чтобы при наличии собственных движений клетки-донора в условиях некогерентного
широкоспектрального облучения реципиента была записана
поляризационно-динамическая голограмма донора.
Во-вторых, этот метод позволяет раскрыть причины виброустойчивости при
регистрации и реконструкции голограмм без лазерных источников света внутри
биосистем в терагерцовом диапазоне волн. Его эффективность определяется
величиной поляризационно-оптической вращательной способности ![]() и толщиной
слоя оптически активной среды
и толщиной
слоя оптически активной среды ![]() . Известно, что вращательная
способность некоторых жидких кристаллов достигает 40000 град/мм, что при
использовании в голографическом информационно-лазерном преобразователе
достаточно для поляризационно-голографической трансляции информации и,
следовательно, голографического управления структурами и процессами в
биосистемах.
. Известно, что вращательная
способность некоторых жидких кристаллов достигает 40000 град/мм, что при
использовании в голографическом информационно-лазерном преобразователе
достаточно для поляризационно-голографической трансляции информации и,
следовательно, голографического управления структурами и процессами в
биосистемах.
Экспериментальные работы по волновому взаимодействию живых систем активно начались с 1980-х гг. Сначала это были работы по взаимодействию клеток [Киркин, 1981; Молчанов, 1985], а затем и по взаимодействию живых организмов [Бурлаков и др., 1999]. Эти работы были успешно продолжены А.В. Будаговским и соавт. [Будаговский, 1990; Будаговский, Евсеева, 1995; Будаговский и др., 1997; Будаговский и др., 2001]. В итоге было показано, что в природе имеет место коммуникационный обмен информацией нехимического (волнового когерентного) происхождения. Такого рода обменные процессы, идущие с участием биорегуляторных сигналов и происходящие без посредства молекулярных и ионных носителей информации, были названы процессами дистанционного межклеточного взаимодействия (ДМВ) [Будаговский, 2004]. Однако казалось невероятным, что слабые электромагнитные клеточные сигналы могут оказывать управляющее влияние на фоне сильных электромагнитных помеховых сигналов природного и техногенного происхождения. Тем не менее, оказалось, что при когерентном приеме световые и другие электромагнитные некогерентные шумы при усреднении обнуляются, тогда, как слабые когерентные и детерминированные сигналы могут накапливаться [Тертышный и др., 1997, 1998, 2000].
В последние годы эти работы получили дальнейшее развитие в Институте проблем управления. В частности, было предложено использовать поляризационно-динамическую голографию, позволяющую сформировать малоподвижные поляризационные кольца. С целью передачи неискаженного изображения каждой точки донора в удаленную зону реципиента был разработан сенсорный квази объектив, а на его основе была создана голографическая установка, предназначенная для экспериментальной проверки возможностей голографического управления.
2. Квантовая телепортация генетико-метаболической информации в пермиссивном варианте (на основе работы [Прангишвили, Гаряев, Максименко и др., 2000(б); http://www.wavegenetics.jino-net.ru/zip/Teleport.zip
Приведенные выше биологические эксперименты по дальнему «переносу» морфогенетической, точнее, генетико-метаболической информации от Донора к Акцептору можно толковать с позиции нелокальных контактов по механизму квантовой телепортации в пермиссивном варианте. Лазерная установка, фактически квантовый биокомпьютер, о которой говорилось выше, кроме уникальной способности к волновому «переносу» морфогенетической информации, осуществляет и конверсию красных когерентных фотонов в радиоволны широкого спектра. Именно они – претенденты на первичные акты считывания-«переноса». При сканировании лазерным лучом биопрепаратов, или любых веществ, по сути осуществляется биологически активная спектроскопия особого рода. Мы предложили предварительное объяснение этого явления [Гаряев, Тертышный, Готовский, 1997], которое является первичным обоснованием нового вида спектроскопии с условным названием – поляризационная лазерно-радиоволновая (ПЛР-спектроскопия) [Прангишвили, Гаряев и др., 2000(б)]. Такая спектроскопия предназначена для исследования неизвестных ранее, вращательно-колебательных квантово-молекулярных характеристик биосистем, биотканей, твердых, жидких, газообразных веществ, а также плазменных состояний. Используемый и описываемый здесь вариант ПЛР-спектроскопии берет узкий оптический диапазон - красный свет, но в дальнейшем планируются модификации с диапазоном от УФ до ИК.
Настоящая версия
установки - это He-Ne лазер (![]() ) с
генерацией двух ортогонально связанных по интенсивности, оптических мод,
которые мобильны, зависят от зондируемого объекта, и связаны таким образом, что
сумма их интенсивностей остается неизменной, независимо от сканируемого
препарата. При взаимодействии хотя бы одной моды с веществом, отраженное или
рассеянное излучение от которого возвращается в оптический резонатор,
происходит перераспределение интенсивности этих оптических мод по закону
изменения поляризации, соответствующей новому состоянию после взаимодействия
луча с динамичными микрополяризаторами, находящимися в сечении освещаемой
площадки исследуемого вещества. Динамичность микрополяризаторов определяется
внутренней динамикой исследуемого объекта (метаболизм, акустика хромосом и ДНК in vitro-in vivo,
колебания кристаллических решеток и т.д.). Одна из мод лазера, в режиме
возврата модулированных фотонов в резонатор, способна в процессе взаимодействия
с веществом быть причиной излучения нашей установкой модулированных радиоволн
широкого спектра, коррелированных с модуляциями в оптических модах излучения
лазера. Эти модуляции зависят от вращательных колебаний микроструктурных
компонентов (например, доменов жидких кристаллов хромосом in vivo-in vitro) исследуемых веществ и их
оптической активности. Частотный интервал радиоволн, образующихся при
конверсии из фотонов, в соответствии с теоретической моделью (см. ниже), лежит
в диапазоне от 2
) с
генерацией двух ортогонально связанных по интенсивности, оптических мод,
которые мобильны, зависят от зондируемого объекта, и связаны таким образом, что
сумма их интенсивностей остается неизменной, независимо от сканируемого
препарата. При взаимодействии хотя бы одной моды с веществом, отраженное или
рассеянное излучение от которого возвращается в оптический резонатор,
происходит перераспределение интенсивности этих оптических мод по закону
изменения поляризации, соответствующей новому состоянию после взаимодействия
луча с динамичными микрополяризаторами, находящимися в сечении освещаемой
площадки исследуемого вещества. Динамичность микрополяризаторов определяется
внутренней динамикой исследуемого объекта (метаболизм, акустика хромосом и ДНК in vitro-in vivo,
колебания кристаллических решеток и т.д.). Одна из мод лазера, в режиме
возврата модулированных фотонов в резонатор, способна в процессе взаимодействия
с веществом быть причиной излучения нашей установкой модулированных радиоволн
широкого спектра, коррелированных с модуляциями в оптических модах излучения
лазера. Эти модуляции зависят от вращательных колебаний микроструктурных
компонентов (например, доменов жидких кристаллов хромосом in vivo-in vitro) исследуемых веществ и их
оптической активности. Частотный интервал радиоволн, образующихся при
конверсии из фотонов, в соответствии с теоретической моделью (см. ниже), лежит
в диапазоне от 2![]() до 0. Максимум такого
радиоизлучения располагается в районе 1 Мгц. Радиоволновой сигнал после
детектирования подается на АЦП компьютера со специальной программой обработки.
На выходе регистрируется Фурье спектр радиоизлучения, характеризующий поляризационно-динамические
свойства изучаемых веществ, с которыми взаимодействует один из лазерных лучей,
а также спектральную память изучаемых веществ. Второй луч при этом возвращается
в резонатор лазера для создания резонансного взаимодействия с атомными
осцилляторами газовой смеси. Причиной конверсии фотонов в радиоволны, как мы
полагаем, является неупругое рассеяние и локализация света основной лазерной
моды на системе неоднородностей зеркал резонатора лазера. Механизм локализации
(локализация в неупругом канале рассеяния) описан ниже. В частности, мы
полагаем, что в резонаторе существует также и упруго рассеянный локализованный
свет. Генерируемое лазером радиоволновое излучение способно «считывать
информацию», например, с препаратов ДНК или органов и тканей. Механизм
«считывания» напоминает механизм обычного индуцированного излучения.
Возможность «открывать и закрывать» лазерный резонатор позволяет локализовать
или «записать» в нем собственные «спектры» различных тестируемых объектов.
Радиоволновое излучение считывает и ретранслирует такие спектры. При этом был
обнаружен эффект спектральной памяти: в течение определенного макроскопического
времени воспроизводятся радиоволновые спектры объектов, отражающих луч обратно
в резонатор и затем удаленных из зоны экспозиции. Так были зарегистрированы
спектры ДНК и выявлена их высокая биологическая активность, вероятно,
связанная с волновым типом «переноса» генетико-метаболической информации
[Гаряев, Леонова, 2003]. Представляется, что квантовая нелокальность генетической
(хромосомной) информации, как проявление ее тотальной распределенности
(континуальности) в пространстве многоклеточных биосистем, является частным
случаем. В действительности, в биосистемах, по крайней мере, шесть уровней
нелокальности.
до 0. Максимум такого
радиоизлучения располагается в районе 1 Мгц. Радиоволновой сигнал после
детектирования подается на АЦП компьютера со специальной программой обработки.
На выходе регистрируется Фурье спектр радиоизлучения, характеризующий поляризационно-динамические
свойства изучаемых веществ, с которыми взаимодействует один из лазерных лучей,
а также спектральную память изучаемых веществ. Второй луч при этом возвращается
в резонатор лазера для создания резонансного взаимодействия с атомными
осцилляторами газовой смеси. Причиной конверсии фотонов в радиоволны, как мы
полагаем, является неупругое рассеяние и локализация света основной лазерной
моды на системе неоднородностей зеркал резонатора лазера. Механизм локализации
(локализация в неупругом канале рассеяния) описан ниже. В частности, мы
полагаем, что в резонаторе существует также и упруго рассеянный локализованный
свет. Генерируемое лазером радиоволновое излучение способно «считывать
информацию», например, с препаратов ДНК или органов и тканей. Механизм
«считывания» напоминает механизм обычного индуцированного излучения.
Возможность «открывать и закрывать» лазерный резонатор позволяет локализовать
или «записать» в нем собственные «спектры» различных тестируемых объектов.
Радиоволновое излучение считывает и ретранслирует такие спектры. При этом был
обнаружен эффект спектральной памяти: в течение определенного макроскопического
времени воспроизводятся радиоволновые спектры объектов, отражающих луч обратно
в резонатор и затем удаленных из зоны экспозиции. Так были зарегистрированы
спектры ДНК и выявлена их высокая биологическая активность, вероятно,
связанная с волновым типом «переноса» генетико-метаболической информации
[Гаряев, Леонова, 2003]. Представляется, что квантовая нелокальность генетической
(хромосомной) информации, как проявление ее тотальной распределенности
(континуальности) в пространстве многоклеточных биосистем, является частным
случаем. В действительности, в биосистемах, по крайней мере, шесть уровней
нелокальности.
1-й уровень – организменный. Нелокальность здесь выражается в способности к регенерации, например у червей планарий. После разрезания таких червей любая часть их тела дает при регенерации целый организм. Иными словами, в этом случае отсутствует привязка общего пула генетической информации к какой-то части биосистемы. То же относится к вегетативному размножению растений.
2-й уровень – клеточный. Из каждой клетки, а не только из зиготы, можно вырастить целый организм. Для животных биосистем это затруднено, но возможно. Каждая клетка – потенциальный континуум организма.
3-й уровень – клеточно-ядерный. Энуклеация ядер из соматических и половых клеток с последующим введением в них других ядер не препятствует развитию нормального организма. Клонирование такого рода уже осуществляют на высших биосистемах, например, на овцах. Каждое клеточное ядро – также потенциальный континуум биосистемы. Локализации генетических потенций на каких-то отдельных клетках нет. Половые клетки выполняют ту же роль, но с гаплоидным набором хромосом, объединяясь в зиготе в двойной набор, как и в соматических.
4-й уровень – молекулярный: рибосома "читает" информационную РНК не только по отдельным кодонам, но и всю ее целиком с учетом контекста, то есть нелокально, континуально.
5-й уровень – хромосомно-голографический. Геном обладает голографической памятью [Gariaev et al., 1991; Гаряев, 1994], а это типично распределенная (нелокальная) ассоциативная память. На этом и последующих уровнях нелокальность приобретает новое качество, дуалистический вещественно-волновой характер, поскольку голограммы как вещество "прочитываются" электромагнитными и/или акустическими полями, выносящими гено-волновую информацию за пределы вещества хромосом. На сцену выходит физическое поле или поля, как калибровочные структуры, размечающие будущее пространство-время организма. Сюда же относится, видимо, голографическая память коры головного мозга, задающая ментальные, смысловые и образные пространства, калибрующие потенциальные действия высших биосистем в социуме как суперорганизме. Это уже высший уровень социо-генетических процессов.
6-й уровень – квантовая нелокальность генома. До 6-го уровня нелокальность генетической информации реализуется в пространстве организма, а также социума. 6-й уровень имеет особый характер и новое качество. Оно проявляется в рамках одной из форм квантовой нелокальности, а именно пермиссивной, постулируемой в данной работе. В этом случае нелокальность мгновенно реализуется как по пространству биосистемы, так и по ее собственному, «сжимаемому» до нуля, времени. Мгновенно распространяемые такими способами гено-волновые программы, изоморфные вещественным, работают в организме "здесь и там одновременно", поэтому утрачивает смысл семантическая конструкция "сначала и потом". И это стратегический фактор, необычайно важное для многоклеточных биосистем эволюционное достижение. Миллиарды клеток организма должны "знать" друг о друге если не все, то очень многое, причем мгновенно. Без явления «волновой информационной мгновенности» гигантский многоклеточный континуум высших биосистем не способен целостно координировать метаболизм, свои физиологические и другие функции. Межклеточная диффузия сигнальных веществ и нервные процессы слишком медленны, инертны для этого. Даже если допустить, что в межклеточной передаче участвуют знаковые электромагнитные поля со световыми скоростями, что достаточно обосновано, то и этого недостаточно. Необходим механизм именно квантовой нелокальности, и он применим к генетическому аппарату, который может выступать как мгновенно распределенный квантовый (волновой) объект, изоморфный вещественным хромосомам. Используя нелокальность, генетический аппарат высших биосистем создает удивительное явление, когда в определенные моменты в «схлопнутом» пространстве-времени биосистемы "здесь и там", "сначала и потом" работают как неразрывность, обеспечивающая организмам суперкогерентность, информационную сверхизбыточность, сверхинформированность, связность и, как итог, целостность (выживаемость). Проявлением этого, например, служит способность к регенерации органов и тканей у низших организмов (гидры, черви, амфибии, ящерицы, ракообразные), способность, которая в значительной степени утрачена человеком. Но ее можно активировать, учитывая развиваемые нами принципы волновой самоорганизации биосистем. Иллюстрацией этого служит первое в мире успешное приживление имплантированных слепому человеку донорских тканей с восстановлением зрения [Мулдашев, 2000]. В основу идеологии такой хирургической операции и регенеративных процессов были положены наши модели морфогенеза и голографической памяти генома. Ключевым моментом данной операции явилось использование донорской сетчатки глаза как голограммы и межклеточного вещества (аллоплант) как системы дополнительной координации постэмбрионального морфогенеза глаза, что было фактически предсказано ранее [Гаряев, 1994].
Вместе с тем, теоретико-экспериментальные исследования здесь все еще носят начальный характер и нуждаются в физико-математическом осмыслении и развитии. Поэтому мы приводим формализованную модель фотонно-радиоволновых процессов, порождаемых при взаимодействии лазерного пучка нашего устройства с веществом, процессов, которые в предположительном плане можно рассматривать как основу ПЛР-спектроскопии и как одну из версий квантовых информационных событий в хромосомах.
В основе наших построений лежат идеи теории локализации света в дисперсных пространственно скоррелированных системах. Явление локализации света получило широкую известность с 1985 года после работы
[Albada, 1985]. Сейчас это одна из наиболее динамично развивающихся областей физики, тесно переплетающаяся с такими проблемами как, например, квантовая телепортация, новые методы записи и считывания информации и т.д.
Рис.5. Схема эксперимента по наблюдению слабой локализации
света.
В работе [Albada, 1985] исследовали отражение света от прозрачной
кюветы, заполненной мельчайшими частицами латекса, взвешенными в воде, в
условиях, когда длина волны падающего фотона ![]() (
(![]() - среднее расстояние
между частицами). На фоне Френелевского отражения, строго в направлении назад
наблюдался очень узкий пик интенсивности рассеянного света (Рис. 5). Сигнал
превышал фоновое значение в 2 раза. Для объяснения эффекта достаточно
рассмотреть рассеяние на паре частиц, оказавшихся на пути фотона. Элемент
траектории фотона, отразившегося в направлении строго назад, есть расположенная
между парой частиц бесконечно узкая петля. Предположим, что эту петлю фотон
может пройти двумя способами - по ходу вращения часовой стрелки и наоборот. Эти
два способа изображены на Рис. 6а). Они неразличимы.
- среднее расстояние
между частицами). На фоне Френелевского отражения, строго в направлении назад
наблюдался очень узкий пик интенсивности рассеянного света (Рис. 5). Сигнал
превышал фоновое значение в 2 раза. Для объяснения эффекта достаточно
рассмотреть рассеяние на паре частиц, оказавшихся на пути фотона. Элемент
траектории фотона, отразившегося в направлении строго назад, есть расположенная
между парой частиц бесконечно узкая петля. Предположим, что эту петлю фотон
может пройти двумя способами - по ходу вращения часовой стрелки и наоборот. Эти
два способа изображены на Рис. 6а). Они неразличимы.
Рис. 6.
а) два способа прохождения фотоном петли на его траектории в условиях слабой
локализации.
б) разворот фотона между двумя частицами.
В
таких случаях квантовая механика предписывает вычислять вероятность Р разворота
фотона следующим образом. Каждому из процессов сопоставляется амплитуда
вероятности а и вероятность разворота ![]() (мы учли, что обе амплитуды под знаком
модуля имеют одинаковые фазы - в этом особенность движения по петле [14]). Если
бы у нас была гипотетическая возможность различить эти способы, вероятность
разворота считалась бы совершенно иначе и была бы в два раза меньше:
(мы учли, что обе амплитуды под знаком
модуля имеют одинаковые фазы - в этом особенность движения по петле [14]). Если
бы у нас была гипотетическая возможность различить эти способы, вероятность
разворота считалась бы совершенно иначе и была бы в два раза меньше: ![]() . Такова
формальная причина пика в направлении назад. Однако появление пика в
направлении назад вовсе не сопровождается соответствующим уменьшением рассеяния
света в каком-либо ином направлении. Как же быть в таком случае с законом
сохранения энергии и откуда взялись добавочные фотоны и почему они не
наблюдаются при отражении света от сплошного полупространства? И третий вопрос.
С чего мы взяли, что существуют два способа движения фотона между парой частиц?
Если траектория фотона между частицами одномерная прямая линия, то о каких двух
различных способах ее обхода может быть речь? Разворот фотона между двумя
рассеивателями - однозначным образом определенная процедура, изображенная на
Рис. 6b.
. Такова
формальная причина пика в направлении назад. Однако появление пика в
направлении назад вовсе не сопровождается соответствующим уменьшением рассеяния
света в каком-либо ином направлении. Как же быть в таком случае с законом
сохранения энергии и откуда взялись добавочные фотоны и почему они не
наблюдаются при отражении света от сплошного полупространства? И третий вопрос.
С чего мы взяли, что существуют два способа движения фотона между парой частиц?
Если траектория фотона между частицами одномерная прямая линия, то о каких двух
различных способах ее обхода может быть речь? Разворот фотона между двумя
рассеивателями - однозначным образом определенная процедура, изображенная на
Рис. 6b.
Итак, нам очень бы хотелось, чтобы было два способа прохождения фотоном бесконечно узкой петли между двумя частицами. Этого можно достичь, если предположить, что топологическая размерность траектории фотона в условиях слабой локализации d<1. Только в этом случае мы можем разместить внутри одной одномерной линии Рис. 6b две разных “линии” - топологический объект, похожий на петлю, т.е. характеризующийся двумя способами его обхода.

Рис. 7. Ожерелье Антуана
Существует изящная математическая конструкция, которая, с одной стороны, очень похожа на то, что в физике называется линией или траекторией, а с другой стороны, ее топологическая размерность d действительно меньше единицы. Более того, d=0. Речь идет о т.н. цепочечном множестве Антуана [Болтянский, Ефремович, 1982]. Этот объект как нельзя лучше приспособлен и для описания процесса непрерывной генерации разномасштабных петель на траектории фотона.
Нульмерное множество Антуана (ожерелье Антуана) устроено следующим образом. На первом этапе рассматривается затравочная “толстая” замкнутая петля А1 . На втором - А1 заменяется цепочкой менее “толстых” звеньев А2 , находящейся внутри А1 . Затем каждое звено А2 заменяется цепочкой еще более мелких звеньев А3 Ì А2 и т.д. Продолжая этот процесс, получим последовательность А1 É А2 É А3 ... (см. Рис. 7). Пересечение этих множеств представляет собой нульмерное антуановское множество А*. Описанная конструкция - простейший вариант антуановского множества.
Несмотря на то, что антуановская цепочка нульмерна, она не утрачивает некоторых свойств обычной одномерной линии. Так, если с обычного нульмерного множества А0 , например, с конечного множества точек легко можно снять “продетое” через него кольцо, нигде не пересекая А0 , то проделать то же самое с нульмерным множеством А* не удается.
Предположим, что траектория фотона в условиях сильной и слабой локализации является антуановским множеством с топологической размерностью d=0. Отсюда следуют интересные выводы. Если фотон движется по антуановской траектории, то покинуть это множество ему довольно трудно. Он испытывает проблемы с выходом в реальный мир с d=1,2,3 , подобные затруднениям человека, находящегося в комнате без окон и дверей. Возможна и физическая интерпретация механизма удержания света в системе, обусловленная необычной топологией антуановских траекторий. Замена реального трехмерного фотона нульмерным объектом приводит к сингулярному характеру распределения энергии вдоль траектории антуановского фотона. У такой траектории появляется своеобразная “механическая жесткость”. Переплетенные “жесткие” звенья антуановского множества сопротивляются любой попытке расцепления. Это также является причиной удержания фотона вблизи пары, точнее, вблизи самого себя.
Рис.8. Антуановские кольца на траектории фотона
Рис.9. Переплетение антуановских колец
Возможен ли выход антуановского фотона в реальный мир? Узкий пик в направлении назад при рассеянии света дисперсной системой в условиях слабой локализации и есть не что иное как испускание антуановских фотонов, инициируемое светом.
Анализ ряда теории возмущений для фотонного пропагатора в системе частиц показывает, что имеются траектории, изоморфные множеству Антуана. Эти траектории, похожие на петлю, составленную из двух частей, как кольцо наручников, представлены на Рис.8. Два полукольца (они не обязательно одинаковые) смыкаются у верхней частицы. Сумма таких петель обозначена нами тонированным кольцом. В процессе своего движения эти кольца траектории могут переплетаться – см. Рис.9. В свою очередь, каждая пропагаторная линия, из которых состоят переплетенные кольца (см. Рис.9), также есть набор переплетенных колец меньшего масштаба (Рис.10) Так повторяется до бесконечности.
Рис.10. Структура пропагаторной линии антуановского кольца
Необходимым
условием для локализации является очень сильная перенормировка или уменьшение
длины волны попадающего в систему фотона. Как известно, в системах с большими
значениями диэлектрической проницаемости длина волны фотона ![]() становится гораздо
меньше длины волны падающего фотона
становится гораздо
меньше длины волны падающего фотона ![]() . Частота фотона
. Частота фотона ![]() при этом не
изменяется - изменяется эффективная скорость фотона
при этом не
изменяется - изменяется эффективная скорость фотона ![]() в соответствии с
соотношением
в соответствии с
соотношением ![]() . Нас интересует ситуация, при которой
. Нас интересует ситуация, при которой ![]() , иначе
фотон не «уместится» на исчезающе малых звеньях антуановского множества.
Эффективная скорость фотона при этом становится нулевой.
, иначе
фотон не «уместится» на исчезающе малых звеньях антуановского множества.
Эффективная скорость фотона при этом становится нулевой.
Одним из объектов, где сильная перенормировка длины волны излучения в действительности возможна, является фрактальный кластер, состоящий из слабо поглощающих частиц-мономеров. Фрактальными называют гетерогенные системы, обладающие масштабной инвариантностью. Любой малый фрагмент системы при увеличении масштаба воспроизводит пространственную структуру всей системы. Фрактальным кластером (ФК) называют обычно агломерат микронных размеров, состоящий из нанометровых твердых частиц, удерживаемых вместе ван-дер-ваальсовскими силами. Фрактальные кластеры образуются либо в результате сильно неравновесной конденсации паров твердого вещества и последующей агрегации нанометровых частиц-мономеров, либо на начальной стадии процесса кристаллизации из растворов или расплавов.
Масштабная инвариантность кластера обусловливает сравнительно медленное спадание парных корреляций в расположении его частиц. Парная корреляционная функция устроена следующим образом
![]()
где D-
фрактальная размерность кластера, ![]() - характерный размер корреляционного
блока. Фрактальная размерность определяет число частиц-мономеров кластера N, находящихся внутри воображаемой сферы радиуса
- характерный размер корреляционного
блока. Фрактальная размерность определяет число частиц-мономеров кластера N, находящихся внутри воображаемой сферы радиуса ![]() :
:![]() . Значение
D<3 и необязательно целое - в этом специфика
фрактального кластера. В обычных плотных упаковках частиц парные корреляции
спадают существенно быстрее, исчезая по экспоненциальному закону на характерных
расстояниях порядка нескольких радиусов частиц. Масштабная инвариантность ФК
визуально отражается в его довольно рыхлой структуре. Плотность частиц в
объеме
. Значение
D<3 и необязательно целое - в этом специфика
фрактального кластера. В обычных плотных упаковках частиц парные корреляции
спадают существенно быстрее, исчезая по экспоненциальному закону на характерных
расстояниях порядка нескольких радиусов частиц. Масштабная инвариантность ФК
визуально отражается в его довольно рыхлой структуре. Плотность частиц в
объеме ![]() фрактального
кластера не постоянна, а пропорциональна
фрактального
кластера не постоянна, а пропорциональна ![]() .
.
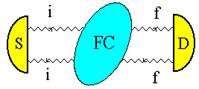
Рис.11. Удержание фотона между источником и детектором при упругом рассеянии на фрактальном кластере
Рис.12. Физические причины удержания фотона
Причиной
перенормировки являются дальнодействующие корреляции в расположении частиц ФК,
визуально выражающиеся в связности кластера и наличии в нем большого числа
пустот. Это объясняется следующим образом. Пусть падающий на кластер фотон с
длиной волны l порядка характерного
размера кластера L улавливается какой-нибудь достаточно крупной полостью ФК
(резонансной полостью). Это улавливание приводит к росту эффективной
диэлектрической проницаемости кластера ![]() (
(![]() растет вблизи любого
электромагнитного резонанса
растет вблизи любого
электромагнитного резонанса
[Борен, Хафмен,
1986]). Возрастание ![]() инициирует, в свою очередь, уменьшение
длины волны фотона, т.к.
инициирует, в свою очередь, уменьшение
длины волны фотона, т.к. ![]() . Фотон с перенормированной длиной
волны
. Фотон с перенормированной длиной
волны ![]() находит
другую полость, меньшего размера. Новое улавливание вновь стимулирует
возрастание
находит
другую полость, меньшего размера. Новое улавливание вновь стимулирует
возрастание ![]() и новое уменьшение
и новое уменьшение ![]() и т.д. В результате
все полости кластера могут оказаться заполненными перенормированными фотонами,
в том числе и теми, чья длина волны
и т.д. В результате
все полости кластера могут оказаться заполненными перенормированными фотонами,
в том числе и теми, чья длина волны ![]() .
.
Физика локализации света в фрактальных системах и схема расчета таковы. Между источником и детектором излучения постоянно присутствует фотон, «циркулирующий» по замкнутой петле (Рис.11). Его удерживает там переплетение жестких антуановских колец на его траектории (Рис.12). Кольца образуются в результате многократного перерассеяния фотона на частицах-мономерах фрактального кластера. В дальнейшем вычисляется амплитуда взаимодействия пары виртуальных фотонов, которые находятся внутри области, обозначенной как FC (фрактальный кластер), на Рис.12. Один из них соответствует верхнему «берегу», второй - нижнему. Типичные процессы, формирующие эту амплитуду можно увидеть на Рис.12, если отбросить волнистые линии реальных фотонов. Амплитуда взаимодействия ищется в виде решения соответствующего уравнения Бете-Солпитера. Можно показать, что мнимая часть этой амплитуды описывает удержание или локализацию фотона в системе.
Соответствующий расчет приводит к следующему выражению для дифференциального сечения упругого рассеяния света кластером [Maksimenko, 1999]:
![]() (1)
(1)
где ![]() ,
, ![]() - угол
рассеяния ,
- угол
рассеяния , ![]() - дельта-функция Дирака, с - скорость
света в вакууме,
- дельта-функция Дирака, с - скорость
света в вакууме, ![]() - единичные векторы поляризации
падающего
- единичные векторы поляризации
падающего ![]() и рассеянного
и рассеянного ![]() квантов,
квантов, ![]() - частота
падающего света и
- частота
падающего света и ![]() - единичный вектор в направлении
рассеянного фотона,
- единичный вектор в направлении
рассеянного фотона, ![]() - число частиц в корреляционном блоке,
- число частиц в корреляционном блоке,
![]() -
диэлектрическая проницаемость материала частиц и
-
диэлектрическая проницаемость материала частиц и ![]() - радиус частицы-мономера. Параметр
- радиус частицы-мономера. Параметр
![]() от
слабо зависит от
от
слабо зависит от![]() . Мнимая часть сечения описывает
«поглощение», обусловленное локализацией. При
. Мнимая часть сечения описывает
«поглощение», обусловленное локализацией. При ![]() это сечение очень велико.
это сечение очень велико.
При Q ¹0
дифференциальное сечение рассеяния становится чисто мнимым. Это означает, что
при Q ¹0
никакого потока рассеянного кластером света вообще не существует. Любой
рассеявшийся “вбок” фотон улавливается кластером и начинает колебаться вдоль
соответствующего ![]() .
.
Не
меньшим сюрпризом выражения (1) для ![]() является сингулярность рассеяния
вперед. Имея в виду связь
является сингулярность рассеяния
вперед. Имея в виду связь
![]()
Рис.13.
Физические причины вынужденного излучения света, локализованного в кластере.
между потоком
рассеянного в направлении ![]() излучения
излучения ![]() и плотностью потока
падающего излучения
и плотностью потока
падающего излучения ![]() , видно, что сингулярность в сечении
означает, что в системе возможен конечный «ток» фотонов даже при нулевой плотности
потока падающего излучения. Сингулярность
, видно, что сингулярность в сечении
означает, что в системе возможен конечный «ток» фотонов даже при нулевой плотности
потока падающего излучения. Сингулярность ![]() в направлении вперед
описывает вынужденное излучение света из кластера. Это типично «лазерный»
эффект. Когерентность вынужденного излучения обеспечивается «нульмерностью»
локализованных антуановских фотонов, способностью к концентрации огромного из
числа в малом объеме. Физическая причина когерентного сброса этих фотонов
проста и наглядна. Любой фотон, рассеявшийся «вбок» улавливается кластером и
начинает осциллировать в нем вдоль направления рассеяния без права выхода из
кластера. На его траектории образуются антуановские кольца, переплетенные с
соответствующими кольцами локализованных фотонов. Именно это переплетение
удерживает такой фотон в кластере. Больше всего таких колец у фотона, рассеивающегося
на нулевой угол - мнимая часть
в направлении вперед
описывает вынужденное излучение света из кластера. Это типично «лазерный»
эффект. Когерентность вынужденного излучения обеспечивается «нульмерностью»
локализованных антуановских фотонов, способностью к концентрации огромного из
числа в малом объеме. Физическая причина когерентного сброса этих фотонов
проста и наглядна. Любой фотон, рассеявшийся «вбок» улавливается кластером и
начинает осциллировать в нем вдоль направления рассеяния без права выхода из
кластера. На его траектории образуются антуановские кольца, переплетенные с
соответствующими кольцами локализованных фотонов. Именно это переплетение
удерживает такой фотон в кластере. Больше всего таких колец у фотона, рассеивающегося
на нулевой угол - мнимая часть ![]() имеет максимум при
имеет максимум при ![]() (см. выражение
(1)). И в то же время только у такого фотона есть возможность вырваться из
кластера, описываемая действительной частью сечения. Этот фотон, зацепившись
своими кольцами за соответствующие кольца локализованных фотонов, вытягивает их
наружу (Рис.13). Так на языке антуановских колец можно легко понять физику
вынужденного излучения света.
(см. выражение
(1)). И в то же время только у такого фотона есть возможность вырваться из
кластера, описываемая действительной частью сечения. Этот фотон, зацепившись
своими кольцами за соответствующие кольца локализованных фотонов, вытягивает их
наружу (Рис.13). Так на языке антуановских колец можно легко понять физику
вынужденного излучения света.
Мы ожидаем, что подобного рода эффекты, а именно - локализация света, имеют место в системе коррелированных зеркал устройства, описанного здесь. Здесь локализация возможна между любой парой из большого числа всевозможных комбинаций зеркал.
Считывание и запись локализованного света
Спектр собственных возбуждений любой системы в немалой степени определяется ее границей или поверхностью. Типичный пример таких возбуждений это плазмон-поляритоны на поверхности металла или поверхностные плазмоны в малых металлических частицах. Существует ли возможность «считать» характерные для такого рода возбуждений спектры и записать их на некотором носителе или накопителе информации с целью, например, длительного хранения и последующего прочтения? Мы расскажем о проблемах и перспективах этих исследований.
Как известно, при отражении фотона от плоской поверхности состояние его поляризации не меняется - это запрещено изотропностью задачи по отношению к вращениям в плоскости поверхности. Казалось бы и при отражении света от плоской пластины с двумя стенками ситуация не изменится. Однако, это не так, если мы учитываем возможность локализации света между границами пластины. Подобного рода эффекты наблюдаются при рассеянии света в направлении строго назад в однородном ансамбле мельчайших частиц
[Максименко и др., 1992]. Связано это с возможностью «вытаскивания» рассеянным назад фотоном фотона, локализованного в системе. В этом случае поляризация отраженного света может измениться. Причина, по которой он «вытаскивает» локализованный фотон, как мы знаем, связана не с фотон-фотонным взаимодействие, которым в данных условиях можно пренебречь, а с переплетением антуановских колец рассеивающегося и локализованного фотонов.
Этот эффект, совмещенный с вращательно-колебательными и поляризационными характеристиками изучаемых объектов, можно использовать для эффективного извлечения из объекта локализованных в нем его собственных возбуждений (его «спектра»). Рассмотрим схему, представленную на Рис.14.
Рис.14. Схема типичного эксперимента по записи ПЛР-спектра исследуемых веществ, например, кристаллических минералов.
В ней фигурирует лазер, описанный выше, и кристалл, чей спектр мы хотим «вытянуть» наружу. В конструкцию стандартного лазера внесено еще одно изменение. Из него удалена полупрозрачная пластинка, расположенная под углом Брюстера к оси лазера (назначение этой пластинки отсекать паразитный свет не основной поляризации). Это делается для того, чтобы не мешать свету, отраженному от кристалла и изменившему свою поляризацию в результате «вытаскивания» из кристалла локализованных фотонов, снова войти в резонатор и затем многократно повторить свой маршрут. Мы ожидаем, что эффективность «вытаскивания» локализованных фотонов, записавших информацию об объекте, в такой системе окажется достаточно высокой для его экспериментального наблюдения. Далее эти делокализованные фотоны могут снова локализоваться но уже в системе зеркал лазера. После этого мы убираем кристалл, но «спектр» его возбуждений, локализованный в лазере, как мы ожидаем, будет еще какое-то время себя проявлять. Система будет воспроизводить спектральную память об объекте, который уже выведен из области экспонирования. Роль кристалла может выполнять любая система, в которой возможна локализация поля. Например, это могут быть биологические объекты, в частности, генетические структуры, которые имеют фрактальную жидкокристаллическую упаковку. Вероятно, именно такого рода эффекты спектральной памяти наблюдались в наших экспериментах (Рис.15).
![]()
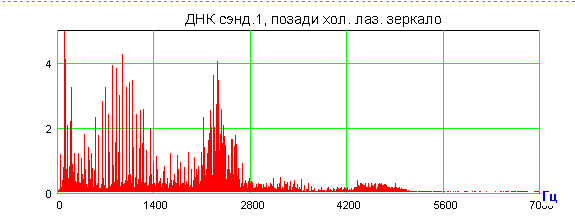
![]()
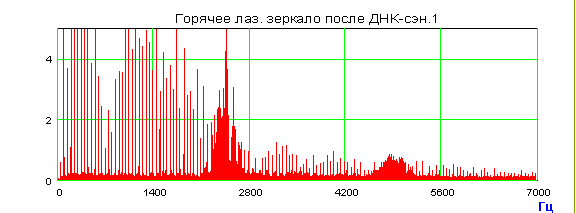
Рис.15.
ПЛР-спектры высокополимерного препарата ДНК из зобной железы теленка (верхний спектр) и его спектрального "следа" на лазерных зеркалах (нижний спектр) после удаления препарата из зоны зондирующего лазерного пучка. Натриевую соль ДНК растворяли в дистиллированной воде в концентрации 1мг/мл, наносили каплю на предметное стекло и закрывали покровным стеклом, получая «стопку». Его экспонировали в лазерном луче в режиме возврата луча, прошедшего через «сэндвич», обратно в резонатор в соответстствии со схемой на Рис.14. Контрольный спектр чистых предметного и покровного стекол в используемом диапазоне частот не дал характерных пиков, типичных для ДНК в составе «стопки».
Подчеркнем,
что речь идет о возможности считывания лазерным излучением с фиксированной
частотой ![]() именно
всего спектра объекта - в широком частотном диапазоне. Дело в том, что
лазерному фотону с частотой
именно
всего спектра объекта - в широком частотном диапазоне. Дело в том, что
лазерному фотону с частотой ![]() «безразлично» какой локализованный
фотон «вытаскивать» из объекта: с той же самой частотой
«безразлично» какой локализованный
фотон «вытаскивать» из объекта: с той же самой частотой ![]() или с любой другой,
если она имеется.
или с любой другой,
если она имеется.
Локализованный свет и проблемы квантовой телепортации.
Совершенно неожиданное приложение идеи локализации света находят в проблеме квантовой телепортации - мгновенной «передаче» послания на произвольно большие расстояния. Эта мощная область исследований, начиная с работ [Bennet et al., 1993; Bouwmeester et al., 1997], привлекает все большее внимание биологов. Коротко напомним основные положения «классической» теории квантовой телепортации.
Как известно,
любая волновая функция пары фотонов (фотон 2 и фотон 3), каждый из которых
обладает двумя состояниями поляризации (горизонтальной поляризацией ![]() и
вертикальной поляризацией
и
вертикальной поляризацией ![]() ), можно разложить по четырем базисным
состояниям (по так называемым состояниям Белла), которые образуют полную
ортонормированную систему функций [22]
), можно разложить по четырем базисным
состояниям (по так называемым состояниям Белла), которые образуют полную
ортонормированную систему функций [22]
![]()
![]()
![]()
![]() (2)
(2)
Состояние ![]() (в
дальнейшем оно нас будет интересовать больше, чем остальные) обладает
интересным свойством: при обнаружении одного из фотонов с определенной
поляризацией поляризация другого оказывается противоположной.
(в
дальнейшем оно нас будет интересовать больше, чем остальные) обладает
интересным свойством: при обнаружении одного из фотонов с определенной
поляризацией поляризация другого оказывается противоположной.
Возможность
экспериментально отличить одно из Белловских состояний от других обеспечивается
их различными симметриями. Из четырех состояний (2) первые три являются
бозонными состояниями (их волновая функция не меняет знак при перестановке
частиц 2 и 3). Последнее состояние ![]() - фермионное (при перестановке 2 и 3
знак волновой функции изменяется). Эта особенность состояния
- фермионное (при перестановке 2 и 3
знак волновой функции изменяется). Эта особенность состояния ![]() позволяет
выделить его в ряде хорошо описанных в литературе экспериментов, использующих
интерференцию двух специальным образом приготовленных световых пучков [Bouwmeester et al., 1997].
позволяет
выделить его в ряде хорошо описанных в литературе экспериментов, использующих
интерференцию двух специальным образом приготовленных световых пучков [Bouwmeester et al., 1997].
Имея в виду
возможность работать далее с состоянием ![]() , в дальнейшем используется ставшая уже
классической следующая экспериментальная схема [Bennet et al., 1993; Bouwmeester et al., 1997; Кадомцев Б.Б., 1999]. Есть два участника игры -
Алиса и Боб, и источник пары фотонов, описываемых состоянием
, в дальнейшем используется ставшая уже
классической следующая экспериментальная схема [Bennet et al., 1993; Bouwmeester et al., 1997; Кадомцев Б.Б., 1999]. Есть два участника игры -
Алиса и Боб, и источник пары фотонов, описываемых состоянием ![]() . Задача
Алисы - передать имеющийся у нее фотон 1 Бобу, находящемуся как угодно далеко
от нее. Однако, Алиса не пользуется обычным классическим способом, а поступает
следующим образом. Алиса и Боб одновременно получают пару фотонов 2 и 3,
описываемую состоянием
. Задача
Алисы - передать имеющийся у нее фотон 1 Бобу, находящемуся как угодно далеко
от нее. Однако, Алиса не пользуется обычным классическим способом, а поступает
следующим образом. Алиса и Боб одновременно получают пару фотонов 2 и 3,
описываемую состоянием ![]() . Алиса получает фотон 2, а Боб - фотон
3. Алиса «смешивает» фотон 1 и 2. При этом в одном случае из четырех она имеет
возможность наблюдать состояние
. Алиса получает фотон 2, а Боб - фотон
3. Алиса «смешивает» фотон 1 и 2. При этом в одном случае из четырех она имеет
возможность наблюдать состояние
![]()
Как только она
это обнаружит, немедленно фотон 3 переходит в начальное состояние фотона 1.
Причина этого следующая. Наблюдение Алисой состояния ![]() означает, что при
каком-то состоянии фотона 1 фотон 2 будет в противоположном по поляризации
состоянии. Но поскольку фотоны 2 и 3 находятся также в состоянии
означает, что при
каком-то состоянии фотона 1 фотон 2 будет в противоположном по поляризации
состоянии. Но поскольку фотоны 2 и 3 находятся также в состоянии ![]() , фотон 3
будет в состоянии, ортогональном состоянию 2, т.е. в состоянии фотона 1. Таким
образом, происходит телепортация фотона 1 от Алисы к Бобу, независимо от
расстояния между ними. Телепортация осуществляется мгновенно.
, фотон 3
будет в состоянии, ортогональном состоянию 2, т.е. в состоянии фотона 1. Таким
образом, происходит телепортация фотона 1 от Алисы к Бобу, независимо от
расстояния между ними. Телепортация осуществляется мгновенно.
Правда, при
такой телепортации поляризационное состояние телепортируемого фотона 1 Алисе
неизвестно, т.к. фотон 1 смешивается с фотоном 2, образуя состояние ![]() .
.
Описанная
процедура телепортации безупречна с точки зрения формализма квантовой механики.
Тем не менее, физический смысл базисных состояний Белла остается неясным, так
же как и нет полной ясности в разрешении парадокса Эйнштейна-Подольского-Розена
(ЭПР-парадокс) [Einstein, Podolsky, Rosen, 1935], для описания которого эти
состояния, собственно, и были введены. Как понять, что при измерении
поляризации ![]() одного из фотонов, находящихся,
например, в состоянии
одного из фотонов, находящихся,
например, в состоянии ![]() , поляризация другого мгновенно
оказывается
, поляризация другого мгновенно
оказывается ![]() , несмотря на то, что их разделяет
очень большое расстояние и любая информация относительно состояния второго
сможет поступить к нам спустя вполне определенный промежуток времени.
, несмотря на то, что их разделяет
очень большое расстояние и любая информация относительно состояния второго
сможет поступить к нам спустя вполне определенный промежуток времени.
Пары фотонов, описываемые состояниями (2) или их линейными комбинациями, называют обычно ЭПР-фотонами или перепутанными фотонами. До тех пор, пока мы не поймем физическую причину мгновенных корреляций в свойствах этих фотонов, мы не поймем физику телепортации, несмотря на всю безупречность логических построений.
Рис.16. Схема эксперимента по считыванию, записи и хранению информации
Как это не удивительно, но к проблемам ЭПР-парадокса и телепортации можно подойти и с другой стороны - с позиций существования локализованного света. Один из вариантов ЭПР-парадокса следующий. Рассматривается, к примеру, s-рассеяние фотона сферической частицей, т.е. рассеянная волна сферически изотропна (см. Рис.16). Пусть рассеянный фотон подходит к детектору в точке А (Алиса). Этот акт регистрации позволяет нам сделать вывод, что в тот же самый момент времени этот рассеянный фотон достигает детектора, расположенного, к примеру, в точке В (Боб), отстоящей от А на сколь-угодно большое расстояние. Это при том, что любая информация из В в А может быть передана по истечении только вполне определенного промежутка времени. Если не рассматривать возможность сверхсветовой скорости распространения сигналов, понять ситуацию можно следующим образом. Что если регистрируемый акт прихода света в А связан вовсе не с рассеянным фотоном, а со сбитым из “трубки” АВ локализованным «длинным» фотоном? Мы “ловим” его левый “конец”. Тогда в том, что в тот же самый момент времени происходит «регистрация» в точке В его “правого” конца, нет ничего странного. Никакого сверхсветового распространения сигнала не происходит, как не происходит и распространения сигнала вообще. «Длинный» локализованный фотон вытаскивается из «полости» за счет зацепления жестких антуановских колец локализованного и рассеивающегося фотонов. Это зацепление аналогично рассмотренному выше зацеплению в фрактальном кластере.
Предположим теперь, что никакого фотона, рассеивающегося на частице нет. А есть «полость» между Алисой и Бобом, заполненная локализованным в ней фотоном. Алиса посылает в эту полость свой фотон. Этот фотон зацепляет по известному нам механизму локализованный фотон и предоставляет его Бобу. Таким образом, в результате действий Алисы, Боб немедленно получает некоторую информацию, правда неизвестно какую, поскольку многие свойства локализованного фотона никому неизвестны.
Как мы видим, в данном случае для мгновенной «передачи» сигнала вместо пары ЭПР-коррелированых фотонов достаточно иметь дело с одним-единственным локализованным фотоном (впрочем, при желании его можно рассматривать как пару взаимодействующих между собой виртуальных фотонов - фотон верхнего берега и фотон нижнего берега рисунков 1 и 2). Кроме того, в [Bouwmeester et al., 1997] ЭПР-пара телепортировала Бобу неизвестный фотон Алисы. В нашем случае фотон Алисы, воздействовав на левый конец никому неизвестного локализованного фотона, предоставляет его правый конец Бобу. В этом вся разница и сходство двух механизмов телепортации.
Противоречит ли телепортация основам специальной теории относительности, утверждающей, что скорость передачи информации не может превосходить скорость света? Очевидно, нет. В случае телепортации беннетовского типа [Bennet et al., 1993; Bouwmeester et al., 1997] мгновенно передается никому неизвестный сигнал. В рамках нашей модели вообще ничего не передается. Боб получает то, что уже находится рядом с ним, но до поры-до времени ему недоступно. Информация уже предсуществует. Алиса мгновенно «разрешает» Бобу взять ее. Поэтому такую модификацию квантовой телепортации (нелокальности) мы назвали пермиссивной (от английского “permission” – разрешение). Надо отметить также, что такая нелокальнсть распространяется, видимо, и дальше, поскольку в нашем случае фотоны, модулированные объектом, мгновенно (нелокально) превращаются в радиоволны, хранящие «фотонную поляризационную информацию». Возможно также, что в наших экспериментах зондирующие объект и интерферирующие встречные фотоны записывают динамичную поляризационную голограмму объекта, например ДНК, и превращают ее в биоактивную радиоволновую, изоморфную фотонной, голограмму.
Генерация радиоволн при неупругом рассеянии света
В этом разделе
мы качественно обсудим одну из возможных причин генерации радиоволн
ПЛР-спектрометром. Речь пойдет о новом механизме неупругого рассеяния света в
электронных системах - в данном случае, в системе металлических слоев
зеркальных покрытий резонатора лазера, являющегося основным элементом
спектрометра. Этот механизм отличается от традиционного комбинационного
рассеяния фотонов. В отличие от дискретного набора стоксовских и
антистоксовских пиков спектр рассматриваемого неупруго рассеянного света
является непрерывным и занимает весь диапазон частот от ![]() до
до ![]() , где
, где ![]() - частота
падающего фотона.
- частота
падающего фотона.
Физика рассматриваемого неупругого рассеяния очень проста. Мы установим основные его закономерности на примере неупругого рассеяния с возбуждением объемных и поверхностных плазмонов в малой металлической частице. Поверхностными плазмонами называют собственные электромагнитные моды мельчайших металлических частиц. Они связаны с собственными колебаниями взаимодействующих через кулоновский потенциал электронов проводимости частицы. Эти моды проявляют себя как отчетливые резонансы в спектрах упругого рассеяния и поглощения света малыми металлическими частицами. Частоты поверхностных плазмонов в зависимости от концентрации электронов проводимости внутри частиц принадлежат границе видимого-УФ света и определяются следующей формулой
![]() ,
,
где ![]() , а w0 - классическая плазменная
частота неограниченного электронного газа
, а w0 - классическая плазменная
частота неограниченного электронного газа
![]() ,
,
где n0 -
плотность электронов проводимости в металле, e и m - заряд и масса электрона.
Возбуждение с ![]() называют дипольным поверхностным
плазмоном, а возбуждение с частотой
называют дипольным поверхностным
плазмоном, а возбуждение с частотой ![]() - объемным плазмоном. Подобного рода
колебания существуют и в тонких металлических пленках, которыми обычно
моделируют зеркальные покрытия, типа тех, что используются в рассматриваемом
лазере. Здесь их называют плазмон-поляритонными модами, их свойства иные, но на
данном этапе мы интересуемся лишь физикой явления.
- объемным плазмоном. Подобного рода
колебания существуют и в тонких металлических пленках, которыми обычно
моделируют зеркальные покрытия, типа тех, что используются в рассматриваемом
лазере. Здесь их называют плазмон-поляритонными модами, их свойства иные, но на
данном этапе мы интересуемся лишь физикой явления.

Рис. 17. Классическая схема неупругого рассеяния фотонов
Классический
механизм неупругого рассеяния света частицей заключается в следующем.
Подлетающий к частице фотон с энергией ![]() возбуждает в ней флуктуацию
электронной плотности, сбрасывая на это часть своей энергии
возбуждает в ней флуктуацию
электронной плотности, сбрасывая на это часть своей энергии ![]() . Энергия
вылетающего фотона
. Энергия
вылетающего фотона ![]() . Этот процесс символически изображен
на Рис. 17. Заштрихованный уголок - флуктуация электронной плотности
. Этот процесс символически изображен
на Рис. 17. Заштрихованный уголок - флуктуация электронной плотности ![]() , которая
есть суперпозиция большого числа электронно-дырочных пар, возбуждаемых фотоном.
Сечение процесса особенно велико, если фотону удается «раскачать» дипольный
поверхностный и объемный плазмоны. Для частицы, чей размер много меньше длины
волны подлетающего фотона, дифференциальное сечение неупругого рассеяния
следующее [Lushnikov et al., 1982]
, которая
есть суперпозиция большого числа электронно-дырочных пар, возбуждаемых фотоном.
Сечение процесса особенно велико, если фотону удается «раскачать» дипольный
поверхностный и объемный плазмоны. Для частицы, чей размер много меньше длины
волны подлетающего фотона, дифференциальное сечение неупругого рассеяния
следующее [Lushnikov et al., 1982]
![]() (1),
(1),
где ![]() - единичный
вектор в направлении рассеянного кванта,
- единичный
вектор в направлении рассеянного кванта, ![]() - угол рассеяния, R - радиус отдельной
частицы пары,
- угол рассеяния, R - радиус отдельной
частицы пары, ![]() и
и ![]() - классический радиус
электрона и комптоновская длина волны электрона соответственно,
- классический радиус
электрона и комптоновская длина волны электрона соответственно,
Если энергии,
сброшенной фотоном, хватит на возбуждение плазмонов ![]() , то
, то
![]()
![]() (2)
(2)
Как мы видим из
анализа выражения (1), возможен только дискретный сброс энергии фотона,
соответствующий возбуждению объемного и дипольного поверхностного плазмона. Это
отражено присутствием дельта-функций Дирака в соответствующем выражении.
Сечение процесса меньше сечения упругого рассеяния света частицей в ![]() раз.
раз.

Рис.18. Предлагаемый механизм неупругого рассеяния фотонов
Предлагаемый
нами механизм принципиально иной. Предположим, что между источником излучения и
детектором непрерывно «циркулирует» по замкнутой петле фотон, многократно
обмениваясь сам с собой флуктуациями электронной плотности, возбуждаемыми в
некоторой системе рассеивателей, находящейся между источником и детектором.
Заштрихованные петли описывают распространение флуктуации электронной плотности
в системе рассеивателей - это так называемые приводимые поляризационные
операторы плотность-плотность или просто корреляторы электронной плотности.
Волнистые линии - волновые функции реальных фотонов, горизонтальные линии -
фотонные пропагаторы. Например, верхняя вершина произвольной нечетной петли
описывает рождение флуктуации электронной плотности фотоном с энергией ![]() за счет
уменьшения его энергии на
за счет
уменьшения его энергии на ![]() , а нижняя - ее схлопывание за счет
получения фотоном энергии
, а нижняя - ее схлопывание за счет
получения фотоном энергии ![]() обратно. Таких петель на траектории
фотона может быть сколько угодно. Наш фотон бесконечное число раз обменивается
энергией сам с собой в процессе неупругого рассеяния. В результате возникает
своеобразное обменное взаимодействие фотона с самим собой, аналогичное обычному
обменному взаимодействию квантовой химии. Именно это взаимодействие удерживает
фотон в «полости» между источником и детектором, обосновывая наше предположение
о возможности столь странной, на первый взгляд, «циркуляции» фотона между
источником и детектором.
обратно. Таких петель на траектории
фотона может быть сколько угодно. Наш фотон бесконечное число раз обменивается
энергией сам с собой в процессе неупругого рассеяния. В результате возникает
своеобразное обменное взаимодействие фотона с самим собой, аналогичное обычному
обменному взаимодействию квантовой химии. Именно это взаимодействие удерживает
фотон в «полости» между источником и детектором, обосновывая наше предположение
о возможности столь странной, на первый взгляд, «циркуляции» фотона между
источником и детектором.
Дифференциальное сечение рассматриваемого процесса имеет вид
![]()
![]()
![]() (3)
(3)
где ![]() и
и ![]() -единичные
вектора поляризации и
-единичные
вектора поляризации и ![]() - сброшенная частота.
- сброшенная частота.
Между
выражениями (2) и (3), несмотря на их внешнее сходство, имеется
принципиальная разница. В рамках классического механизма возможен только
дискретный сброс энергии падающего фотона, соответствующий возбуждению
объемного (с частотой ![]() ) и дипольного поверхностного плазмонов
(частота
) и дипольного поверхностного плазмонов
(частота ![]() )
в частицах (любой другой сброс энергии запрещен фигурирующими в (1)
)
в частицах (любой другой сброс энергии запрещен фигурирующими в (1) ![]() -функциями).
Что касается предлагаемого механизма, то красный сдвиг частоты падающего фотона
может быть любым в интервале. Если
-функциями).
Что касается предлагаемого механизма, то красный сдвиг частоты падающего фотона
может быть любым в интервале. Если ![]() , результатом процесса является
наблюдаемая экспериментально генерация радиоволн.
, результатом процесса является
наблюдаемая экспериментально генерация радиоволн.
Наряду
с «красным» смещением возможен и «голубой» сдвиг частоты фотона.. Таким
образом, спектр неупруго рассеянного света с учетом локализации должен занимать
весь диапазон частот от 0 до ![]() . Подобного рода эффекты действительно
наблюдаются в экспериментах по гигантскому комбинационному рассеянию света
молекулами, адсорбированными на поверхности мельчайших металлических частиц -
его называют «гигантский белый фон», и он до сих пор остается загадкой.
. Подобного рода эффекты действительно
наблюдаются в экспериментах по гигантскому комбинационному рассеянию света
молекулами, адсорбированными на поверхности мельчайших металлических частиц -
его называют «гигантский белый фон», и он до сих пор остается загадкой.
Процессы Рис. 17
при ![]() качественно
объясняют повышенный фон радиоизлучения рассматриваемого лазера. Количественный
расчет безусловно требует учета специфики системы.
качественно
объясняют повышенный фон радиоизлучения рассматриваемого лазера. Количественный
расчет безусловно требует учета специфики системы.
3. Молекулярно-тканевые «радиостанции».
Дополнительные теоретические подходы здесь, возможно, лежат в эффектах так называемого «слабого воздействия» [Чукова, 2002]. Видимо, регенерационные и цитопротекторные эффекты в наших экспериментах носят эндоэргический характер, когда даже слабо поглощаемая биопрепаратами энергия когерентного красного поляризованного лазерного излучения идет на увеличение свободной энергии Гельмгольца, аккумулированной в химических связях метаболитов биопрепаратов, что приводит к существенным биоэффектам. Атомы информационных макромолекул (ДНК, РНК, белков), взаимодействуя с лазерным лучом, вместе с энергией квантов света приобретают и их момент количества движения, что создает инверсную заселенность ядерных зеемановских уровней, т.е. происходит так называемая химическая поляризация ядер. Квантовый выход поляризации, т. е. число избыточных ядерных спинов на верхнем зеемановском уровне, приходящееся на каждый поглощенный квант света, может составлять 30%. Инверсно заселенная протонно-спиновая система может испускать кванты с энергией порядка 6,5∙10-26 Дж, которые соответствуют частотам около 100 МГц [Бучаченко, 1979].
Развивая и детализируя сказанное, можно думать, что при сканировании биопрепаратов лазерным светом возникают комбинационные частоты, захватывающие синий и УФ диапазон. Кроме того, как мы ранее предлагали в соответствующей модели локализованного света, происходит преобразование частот в диапазоне от 2 омега до нуля [Прангишвили, Гаряев и др., 2000(б)]. И кроме того, при сканировании биопрепаратов имеет место широкополосное радиоизлучение газового разряда лазера. Учитывая это, полагаем, что в наших экспериментах биопрепараты-доноры сканируются («считываются» лазером) многочастотным путем. Метаболизирующие биохимические составляющие биопрепаратов, которые взаимодействуют с динамически поляризованным красным светом зондирующего лазера (и широким спектром дополнительных излучений), могут генерировать электромагнитные радиочастотные колебания. В этой ситуации препараты поджелудочной железы и селезенки выступают в роли своеобразных органно-молекулярных радиостанций, где каждый вид их молекул имеет свои характерные частоты, которые могут усиливаться благодаря стохастическим резонансам. С другой стороны, определенные типы молекул пораженной аллоксаном поджелудочной железы больных животных и/или стволовые клетки их крови могут резонансно поглощать такое радиоизлучение, которое несет информационную составляющую для запуска биохимических реакций, которые приводят к регенерации тканей поджелудочной железы и защитным антиаллоксановым процессам. Это не исключает существенной роли ранее обсуждавшихся механизмов, связанных с телепортационными и голографическими характеристиками генома биопрепаратов-доноров. Обратимся к известным положениям химии и физики http://www.chem.msu.su/rus/publ/Buchachenko/buch5.html, задающим квантовый сценарий рассматриваемых генетико-биохимических событий. В ансамбле молекул-продуктов с инверсной населенностью в зеемановском резервуаре запасается энергия; она может растратиться в тепло (через спин-решеточную магнитную релаксацию), но может превратиться в стимулированное излучение на зеемановской ядерной частоте. В этом случае реакция действительно становится радиочастотным эмиттером, квантовым генератором с химической накачкой (подобно химическим лазерам). Это новое явление - радиоизлучение химической реакции - сначала было предсказано теоретически, а затем обнаружено экспериментально. Оно возникает, когда энергия зеемановского резервуара превосходит порог генерации; тогда движение ядерных спинов спонтанно становится когерентным, и такая когерентная система ядер становится квантовым генератором, излучающим в микроволновой области. Но это лишь одна сторона химической радиофизики. Химическая реакция может быть не только генератором, но и приемником микроволн. Прием на химическом уровне следует из принципов спиновой химии: резонансное микроволновое излучение стимулирует триплет-синглетную конверсию радикальных пар (или пар других спиновых носителей) и изменяет выход химических продуктов. Таким образом магнитно-спиновые эффекты делают биохимическую реакцию химическим приемником микроволнового излучения. Более того, такой прием можно осуществлять селективно. Если микроволновая накачка затрагивает все радикальные пары (биохимических субстратов), то общий результат сводится к изменению выхода продуктов реакции на резонансных частотах. Этот эффект назван магнитным резонансом, детектируемым по выходу продуктов реакции - РИДМР (от английского reaction yield detected magnetic resonance, RYDMR). Если накачка селективна и затрагивает только радикальные пары с магнитными ядрами, возникает явление - радиоиндуцированный магнитный изотопный эффект (РИМИЭ). И, наконец, если микроволновая накачка селективна еще и по ориентации ядерных спинов (т.е. затрагивает ансамбли радикальных пар с избранной ориентацией ядерных спинов), то появляется стимулированная поляризация ядер (СПЯ). Это связано с т.н. спиновым катализом. Он примечателен тем, что спиновая конверсия реагентов индуцируется парамагнитной частицей - спиновым катализатором. Конверсия происходит в результате обменного взаимодействия катализатора (фермента) с реагентами. Спиновый катализ ускоряет рекомбинацию радикалов, уакс-изомеризацию соединений с двойной связью (на семь-восемь порядков), рекомбинацию спин-поляризованных атомов и т. д. Возможно, что спиновый катализ действует и в биохимических процессах указанного процесса регенерации поджелудочной железы и в цитопротекторном эффекте. В основе спиновой химии и химической радиофизики лежит манипулирование электронными и ядерными спинами. Когда такие манипуляции производит сама химическая реакция, появляются магнитно-спиновые эффекты, и в их числе - генерация микроволн, когда реакция становится молекулярной радиостанцией. Когда манипуляции со спинами осуществляются под воздействием микроволн, рождаются и другие магнитные эффекты второго поколения. Они служат индикатором приема микроволн. И спиновая химия, и химическая радиофизика тесно связаны, но у них есть и самостоятельные задачи. Первая разрабатывает новые принципы управления химическими реакциями (в том числе и с помощью микроволн), вторая имеет крупный прикладной биомедицинский аспект.
В связи с версией молекулярно-тканевых «радиостанций» возникает принципиальный вопрос о причине высокой проникающей способности мШЭИ. Как мы помним, одна из групп крыс в наших экспериментах была помещена в изолированное бетоном подвальное помещение, и тем не менее, эффект воздействия мШЭИ на животных была достоверн зафиксирован. Если биологически (морфогенетически) действующая часть мШЭИ занимает микроволновый диапазон зеемановского резервуара, то этот участок спектра мШЭИ будет отфильтрован бетонными перекрытиями подвального помещения, где находились крысы в момент волнового воздействия на них. Но крысы воздействие воспринимают. Каким образом? Возможное объяснение состоит в следующем. Электроны зеемановских уровней энергий всех метаболитов, включая генетические структуры, находясь в потенциальной "энергетической яме", испытывают сверхтонкий лэмбовский сдвиг этих уровней порядка 1000 МГц. Он обусловлен существованием продольных фотонов ядер атомов, создающих его продольное (электростатическое) поле, которое дипольно возмущает вакуум и с этим возмущением взаимодействуют двигающиеся орбитальные электроны атома. В свою очередь, эти электроны имеют собственное электростатическое поле, состоящее из аналогичных продольных фотонов. Таким образом, атомная система электронов (переменная во времени) индуцирует вокруг себя составное переменное продольное электрическое поле, которое в форме продольной электрической волны (ПЭВ) двигается мгновенно во всём окружающем пространстве до бесконечности. Вектор Умова-Пойтинга этой волны равен нулю, т.е. энергия-импульс данной атомной системой не излучается. Однако существуют вихри продольного электрического поля Максвелла, описываемого вещественной частью бикватернионов [Березин и др., 2003], которые могут переносить информацию, имеющую численный энергетический эквивалент Бриллюэна-Шеннона. Полученная таким путем ПЭВ, обладая аномально высокой проникающей способностью, проходит почти без затухания через различные препятствия (металлические экраны, ферромагнетики и диэлектрики, бетон). Ядра клетки и их основной компонент – ДНК – возбуждают солитоны [Смелов, 2001] связанных волн электронов и гиперзвуковых колебаний жидкокристаллических хромосомных структур, т.е. электрон-вибронные колебания [Берсукер, 1976] или электрон-ядерные волновые колебания двойной спирали ДНК. Указанные электрон-вибронные волны переизлучают (рассеивают) падающую ПЭВ обратно в эфир и могут приниматься другими биосистемами подобными той, на которую воздействует первичная волна возбуждения ПЭВ.
В силу высокой добротности ~1014 всех электрон-вибронных колебательных систем, они обладают высокой чувствительностью, исчисляемой долями планковского кванта энергии в расчёте на один элемент когерентной колебательной цепочки, которая, например, может быть спиралью ДНК или мембраной клетки. В открытых состояниях спирали ДНК, инициируемых тепловым движением в живой клетке, электрон-вибронные колебания существуют в виде солитонных (геликоидальных, винтовых, вихревых) движений атомов цепочки. Это так называемые солитоны Салерно-Маслова, и они способны к считыванию информации с «текстов» ДНК-РНК последовательностей. Излучение таких «информационных» солитонов генерируются электрон-вибронными колебаниями ДНК и РНК. При этом считанная с генетических молекул информация может передаваться другим клеткам (и за пределы биосистем) эстафетно по механизму рассеяния с изменением частот ПЭВ, излученных в область радиочастот и акустических частот. Причём, информационное радиоизлучение электрон-вибронных колебаний в форме ПЭВ на определённых частотах, в принципе, может управлять биохимическими процессами. И наоборот, биохимические реакции в препаратах, зондируемых поляризованным лазерным излучением, могут генерировать электромагнитные радиочастотные колебания.
В связи с вероятной волновой генетико-знаковой нагрузкой вихревых солитонных состояний, инициируемых на молекулах ДНК и РНК in vivo, рассмотрим эти процессы более детально [Благодатских, Гаряев и др., 1996; Гаряев, 1997].
Математическое моделирование солитонов на ДНК
Марио Салерно первым начал компьютерное экспериментирование с солитонами на ДНК не только как с формальными математическими структурами, он попытался связать их поведение в одномерном пространстве полинуклеотидов с их биогенетическими, а точнее, с эпигенетическими функциями. При этом он развил первую модель солитонов на ДНК, предложенную Инглендером и соавторами. Эта модель и в последующем ее более детальные формы, включая нашу (см. ниже), представлена в понятиях механических систем как цепочка осцилляторов (оснований ДНК), связанных упругими нелинейными сахаро-фосфатными связями. Вслед за Салерно основное внимание мы уделили реально существующим известным последовательностям ДНК и влиянию их на характер поведения солитонов. На первом этапе мы повторили его эксперименты, но на существенно более длинных отрезках ДНК. Действительно, солитонные возбуждения типа кинков чувствительны к месту своей инициации, и продвижение их вдоль одной из цепочек ДНК, когда они раскрыты вследствие тепловых флуктуаций, сопровождается специфической модуляцией траектории кинков во времени и в одномерном пространстве полинуклеотидов. Такие солитоны являются структурами, излучающими электромагнитное и акустическое поле, их внутренняя колебательная структура способна отобразить и ретранслировать тексты и иные знаковые структуры ДНК во внутри- и внеклеточное пространство, по крайней мере, на уровне крупных блоков последовательностей. В качестве примера можно привести поведение кинка на фрагменте одиночной цепи ДНК длиной 1020 нуклеотидов из вируса саркомы птиц.
C-район ДНК (1 ![]() 1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит
несколько “семантически” определенных участков, таких, как
полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) - 3’-конец вирусной РНК, сайт
полиаденилирования; 916 нуклеотид - 5’-конец вирусной РНК (“capping site”); Red-участок
(917 - 936) - короткий концевой повтор вирусного
генома; Pro - вероятный компонент промотора
транскрипции (между 870 и 900); палиндром-”шпилька” (870 - 912)[2].
1020 нуклеотид) на 3’-конце вируса саркомы птиц. Содержит
несколько “семантически” определенных участков, таких, как
полипептид-кодирующий участок (между 558 и 675 нуклеотидами); PolA (936) - 3’-конец вирусной РНК, сайт
полиаденилирования; 916 нуклеотид - 5’-конец вирусной РНК (“capping site”); Red-участок
(917 - 936) - короткий концевой повтор вирусного
генома; Pro - вероятный компонент промотора
транскрипции (между 870 и 900); палиндром-”шпилька” (870 - 912)[2].
(5’- начало) ![]() GGC CTA TGT GGA GAG GAT GAA CTA CGT GCA CCG AGA CCT GCG GGC GGC CAA CAT CCT GGT GGG GGA GAA CCT GGT GTG CAA GGT GGC TGA CTT TGG GCT GGC ACG CCT CAT CGA GGA CAA CGA GTA CAC AGC ACG GCA AGG TGC AAG TTC CCC ATC AAG TGG AGA GCC CCC GAG GCA GCC CTC TAT GGC CGG TTC ACC ATC AAG TCG GAT GTC TGG TCC TTC GGC ATC CTG CTG ACT GAG CTG ACC ACC AAG GGC CGG GTG CCA TAC CCA GGG ATG GGC AAC GGG GAG GTG CTG GAC CGG GTG GAG AGG GGC TAC CGC ATG CCC TGC CCG CCC GAG TGC CCC GAG TCG CTG CAT GAC CTT ATG TGC CAG TGC TGG CGG AGG GAC CCT GGA GGA GCG GCC CAC TTT TCG AGC TAC CTG CAG GCC CAG CTG CTC CCT GCT TGT GTG TTG GAG GTC GCT GAG TAG TGC GCG AGT AAA ATT TAA GCT ACA ACA AGG CAA GGC TTG ACC GAC AAT TGC ATG AAG AAT CTG CTT AGG GTT AGG CGT TTT GCG CTG CTT CGC GAT GTA CGGGCC AGA TAT ACG CGT ATC TGA GGG GAC TAG GGT GTG TTT AGG CGA AAA GCG GGG CTT CGG TTG TAC GCG GTT AGG AGT CCC CTC AGG ATA TAG TAG TTT CGC TTT TGC ATA GGG AGG GGG AAA TGT AGT CTT ATG CAA TAC TCT TGT AGT CTT GCA ACA TGG TAA CGA TGA GTT AGC AAC ATA CCT TAC AAG GAG AGA AAA AGC ACC GTG CAT GCC GAT TGG TGG AAG TAA GGT GTA CGA TCG TGC CTT ATT AGG AAG GCA ACA GAC CGG GTC TGA CAT GGA TTG GAC GAA CCA CTG AAT TCC GCA TCG CAG AGA TAT TGT ATT TAA GTG CCT AGC TCG ATA CAA TAA ACG CCA TTT GAC CAT TCA CCA CAT TGG TGT GCA CCT GGG TTG ATG GCT GGA CCG TCG ATT CCC TAA CGA TTG CGA ACA CCT GAA TGA AGC AGA AGG CTT CATT
GGC CTA TGT GGA GAG GAT GAA CTA CGT GCA CCG AGA CCT GCG GGC GGC CAA CAT CCT GGT GGG GGA GAA CCT GGT GTG CAA GGT GGC TGA CTT TGG GCT GGC ACG CCT CAT CGA GGA CAA CGA GTA CAC AGC ACG GCA AGG TGC AAG TTC CCC ATC AAG TGG AGA GCC CCC GAG GCA GCC CTC TAT GGC CGG TTC ACC ATC AAG TCG GAT GTC TGG TCC TTC GGC ATC CTG CTG ACT GAG CTG ACC ACC AAG GGC CGG GTG CCA TAC CCA GGG ATG GGC AAC GGG GAG GTG CTG GAC CGG GTG GAG AGG GGC TAC CGC ATG CCC TGC CCG CCC GAG TGC CCC GAG TCG CTG CAT GAC CTT ATG TGC CAG TGC TGG CGG AGG GAC CCT GGA GGA GCG GCC CAC TTT TCG AGC TAC CTG CAG GCC CAG CTG CTC CCT GCT TGT GTG TTG GAG GTC GCT GAG TAG TGC GCG AGT AAA ATT TAA GCT ACA ACA AGG CAA GGC TTG ACC GAC AAT TGC ATG AAG AAT CTG CTT AGG GTT AGG CGT TTT GCG CTG CTT CGC GAT GTA CGGGCC AGA TAT ACG CGT ATC TGA GGG GAC TAG GGT GTG TTT AGG CGA AAA GCG GGG CTT CGG TTG TAC GCG GTT AGG AGT CCC CTC AGG ATA TAG TAG TTT CGC TTT TGC ATA GGG AGG GGG AAA TGT AGT CTT ATG CAA TAC TCT TGT AGT CTT GCA ACA TGG TAA CGA TGA GTT AGC AAC ATA CCT TAC AAG GAG AGA AAA AGC ACC GTG CAT GCC GAT TGG TGG AAG TAA GGT GTA CGA TCG TGC CTT ATT AGG AAG GCA ACA GAC CGG GTC TGA CAT GGA TTG GAC GAA CCA CTG AAT TCC GCA TCG CAG AGA TAT TGT ATT TAA GTG CCT AGC TCG ATA CAA TAA ACG CCA TTT GAC CAT TCA CCA CAT TGG TGT GCA CCT GGG TTG ATG GCT GGA CCG TCG ATT CCC TAA CGA TTG CGA ACA CCT GAA TGA AGC AGA AGG CTT CATT ![]() 1020 (3’-конец)
1020 (3’-конец)
На Рис.1 и 2 кинки имеют форму “горных гряд”, а не ступенек, поскольку взята производная от функции уравнения синус-Гордона. Здесь горизонтальная ось - последовательность ДНК, вертикальная - амплитуда солитона. Ось на зрителя - время. Видно, как при изменении места инициации солитона на определенных последовательностях полинуклеотида заметно меняется динамика этой уединенной волны в форме ее вращательно-колебательных движений вдоль цепочки ДНК.
Исследуемый район молекулы богат функционально (семантически) биологически значимыми участками, и мы вправе ожидать, что они, эти участки, будут изменять, модулировать, то есть вводить в одиночную цепочку ДНК или в РНК “текстовую” информацию. Таковая будет реализоваться в спектре колебаний солитонной волны вдоль цепочки полинуклеотида. Такой спектр будет отображать последовательности нуклеотидов и, тем самым, будет исполнять роль переносчика генетических сообщений. Такая модуляция колебательной структуры солитонов отчетливо наблюдается на приведенных графиках. Можно полагать, что спектральный состав частот колебаний солитонов является одним из механизмов преобразования текстовых структур ДНК и РНК в волновую форму и средством передачи генетических и иных сообщений в одномерном пространстве вдоль цепочек полинуклеотидов и (или) в трехмерном измерении генома как отдельной клетки, так и тканевого континуума биосистемы.
x z y
![]()
![]()
![]()

400-ый
Рис.1
Влияние нуклеотидной последовательности ДНК на динамику конформационного возмущения волны солитона. Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 400-ый нуклеотид.
y – амплитуда солитона, x – длина полинуклеотида (их число), z – время.

450-ый
Рис.2
То же, что на Рис.1, но центр возмущения цепочки ДНК на 450-ом нуклеотиде.
Так работает компьютерная модель динамики солитонов, в определенной мере развитая Салерно после ее выдвижения Инглендером. Салерно дал формализм, описывающий вращательные колебания нуклеотидов молекулы ДНК, для того чтобы объяснить экспериментальные данные по водородно-тритиевому обмену в ДНК. Согласно этой модели, по Инглендеру, в цепи ДНК могут возникать (под воздействием теплового шума) и распространяться открытые состояния (“плавление” двойной спирали ДНК на коротких участках, обогащенных АТ-парами ) в виде локализованных дислокаций.
Марио Салерно, продолжая работу Инглендера в упрощенном варианте, выявил влияние последовательности нуклеотидов на нелинейную солитонную динамику вращательных колебаний нуклеотидов на однотяжных участках ДНК, образующих такие открытые (“open state”) области. Позднее Якушевич, Федянин, Хомма и др. рассмотрели различные обобщения модели Инглендера, с оценкой особенностей строения ДНК, учитывая обрыв водородной связи при открытии оснований, парность цепи ДНК и другие степени свободы, отличные от вращательных. Однако в указанных работах недостаточно сказано о причинах возникновения дислокаций в ДНК. Мы предлагаем возможный механизм этого процесса в ДНК, альтернативный гипотезе Инглендера о воздействии теплового шума как причины раскрытия пар оснований. Мы считаем, что дислокации на ДНК могут возникать при изменении периода спирали ДНК (основная часть идеи принадлежит М.Ю.Маслову). В нашей модели нуклеотиды ДНК рассматриваются как осцилляторы, подвешенные на невесомом нерастяжимом стержне; сахаро-фосфатная связь между соседними нуклеотидами в цепи моделируется линейными пружинами; спирализация вдоль цепи не учитывается; водородные связи между комплементарными основаниями моделируется “гравитационным” потенциалом. Гамильтониан по М. Салерно выглядит следующим образом:
![]() (1)
(1)
где: ![]() - углы вращений нуклеотидов в разных
цепях,
- углы вращений нуклеотидов в разных
цепях, ![]() - константы упругости вдоль цепей,
- константы упругости вдоль цепей, ![]() - число пар в цепи,
- число пар в цепи, ![]() - момент инерции оснований,
- момент инерции оснований, ![]() - константа упругости водородных
связей между комплементарными основаниями.
- константа упругости водородных
связей между комплементарными основаниями.
Коэффициенты ![]() в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом: ![]() в случае АТ и ТА пар,
в случае АТ и ТА пар, ![]() в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар; ![]() - параметр, определенный Федяниным и Якушевич[3] и полученный на основе модели
синус-Гордона и экспериментальных данных. Далее для упрощения модели считается,
что
- параметр, определенный Федяниным и Якушевич[3] и полученный на основе модели
синус-Гордона и экспериментальных данных. Далее для упрощения модели считается,
что ![]()
Уравнения
движения для разности ![]() , полученные из (1), имеют по М. Салерно вид:
, полученные из (1), имеют по М. Салерно вид:
![]() (2)
(2)
где
произведена замена ![]() .
.
В случае ![]() , в системе (2) можно перейти к безразмерному
дифференциальному уравнению синус-Гордона:
, в системе (2) можно перейти к безразмерному
дифференциальному уравнению синус-Гордона:
![]() ,
(3)
,
(3)
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, соответствует дислокации в цепи.
Основным
предположением моделей Инглендера-Салерно является то, что взаимодействие между
комплементарными основаниями описывается потенциалом ![]() (4), в котором не учитывается обрыв водородной связи.
(4), в котором не учитывается обрыв водородной связи.
В нашей работе рассматривается следующий потенциал:
![]()
Кроме того,
учитывается вязкость водной среды (в воде вязкость ![]() ~ 1).
~ 1).
Рассматриваются также факторы, приводящие к спирализации ДНК, при этом они считаются внешними силами, задаваемыми потенциалом

где
![]() -
период спирали.
-
период спирали.
Уравнения
(2) с потенциалом ![]() и с учетом вязкости
принимают вид:
и с учетом вязкости
принимают вид:
![]() (5)
(5)
Известно,
что период спирали ДНК меняется в зависимости от влажности. В частности, для
кристаллической ДНК ![]() , а в водной среде
, а в водной среде ![]() -
в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление
суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или
замкнутыми концами) возникает напряжение, связанное с недостатком (избытком)
количества витков спирали до релаксированного состояния. Если
-
в пределах от 10. 3 до 10. 6. Именно этим фактором обусловлено явление
суперспирализации. При изменении шага спирали в цепи ДНК (с фиксированными или
замкнутыми концами) возникает напряжение, связанное с недостатком (избытком)
количества витков спирали до релаксированного состояния. Если ![]() ,
то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар
оснований возникнет избыток в
,
то при переходе из сухого в увлажненное состояние для цепи длиной в 300 пар
оснований возникнет избыток в ![]() витка.
витка.
На основе результатов численного моделирования, представленных ниже, предполагается, что изменение шага спирали может привести не только к суперспирализации, но и к локальному расхождению обеих комплементарных цепочек ДНК. Кроме того, при суперспирализации напряжение в цепи снимается не полностью, поэтому локальное расхождение, вероятно, может происходить и одновременно с суперспирализацией.
Система
(5) численно интегрировалась в интервале ![]() с шагом
с шагом ![]() .
Начальные условия следующие:
.
Начальные условия следующие:
![]()
![]()
![]()
Период
спирали в системе (5)![]() длина poly(A)-цепи -
300 пар оснований. То есть параметры периода спирали в начальных условиях и в
системе (5) различны. Таким образом, моделирован перенос ДНК из
кристаллического состояния в увлажненное, близкое состоянию in
vivo.
длина poly(A)-цепи -
300 пар оснований. То есть параметры периода спирали в начальных условиях и в
системе (5) различны. Таким образом, моделирован перенос ДНК из
кристаллического состояния в увлажненное, близкое состоянию in
vivo.
Граничные условия следующие (назовем их “квазициклическими”):
![]()
Особенностью данной модели является то, что при переходе из состояния с периодом в 10 пар в состояние с периодом в 10, 5 пар почти вся цепь оказывается денатурированной (“расплавленной”). Приведенные ниже результаты описывают процесс ренатурации такой цепи с возникновением дислокаций.
В этих
экспериментах варьировали параметры: 1) диссипация ![]() 2) отношение параметров упругости
2) отношение параметров упругости ![]() 3) угол обрыва водородных связей
3) угол обрыва водородных связей ![]() .
.
На Рис. 3 и 4
представлены результаты численного интегрирования системы (5). Показана не сама
функция ![]() , а разница
, а разница ![]() , поскольку область изменения функции
, поскольку область изменения функции ![]() (приблизительно от
(приблизительно от ![]() до
до ![]() ) велика по сравнению с характерными изменениями в системе
(приблизительно от 0 до 9). Горизонтальная часть графиков соответствует не
разошедшемуся участку ДНК (двойная цепочка) с периодом спирали
) велика по сравнению с характерными изменениями в системе
(приблизительно от 0 до 9). Горизонтальная часть графиков соответствует не
разошедшемуся участку ДНК (двойная цепочка) с периодом спирали ![]() . Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации.
. Наклонная часть графиков на рис. 3(a), 4(а) соответствует дислокации.
Исходя из этой модели, можно полагать, что
1)
Способность к образованию дислокации в этой модели сильно зависит от ![]() . При
. При ![]() дислокация возникла во всех рассмотренных случаях.
дислокация возникла во всех рассмотренных случаях.
2)
Способность к образованию дислокации также сильно зависит от параметра![]() . Во всех случаях, когда параметр
. Во всех случаях, когда параметр ![]() велик (
велик (![]() ),
),
на Рис. 1.а, 2.а ), дислокация возникла. В пользу этого свидетельствует также сравнение Рис. 3(а) и 4(г).
Как
показывают дополнительные расчеты, влияние![]() на эффект проявляется в меньшей степени. Дислокация
образуется или не образуется вне зависимости от значения
на эффект проявляется в меньшей степени. Дислокация
образуется или не образуется вне зависимости от значения ![]() (
(![]() или
или ![]() ). При больших значениях
). При больших значениях ![]() дислокация образуется медленнее, чем при меньших.
дислокация образуется медленнее, чем при меньших.
3) На Рис. 3(а), 4(в,г) видно, что дислокация имеет кинкообразную форму.
Ширина
дислокации зависит от параметров ![]() (чем больше
(чем больше ![]() , тем меньше ширина дислокации) и
, тем меньше ширина дислокации) и ![]() (чем больше
(чем больше ![]() , тем меньше ширина дислокации).
, тем меньше ширина дислокации).
Развивая дальше модели солитонных возбуждений в ДНК (совместно с М.Ю.Масловым и др.) мы использовали условия, при которых цепочки ДНК моделируются набором ровибронных осцилляторов, подвешенных на невесомом нерастяжимом стержне; для простоты спирализация цепи не учитывается, а ровибронные степени свободы одной из цепочек считаются “замороженными”.
В этом случае гамильтониан для “активной” цепочки записывается в следующем виде:
H=H0+H1+H2
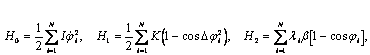 (1)
(1)
где: ![]() - число пар оснований в цепи;
- число пар оснований в цепи; ![]() - гамильтониан, описывающий собственные
осцилляции мономеров (
- гамильтониан, описывающий собственные
осцилляции мономеров (![]() - углы вращения нуклеотидов в цепочке,
- углы вращения нуклеотидов в цепочке,![]() - момент инерции оснований);
- момент инерции оснований); ![]() - гамильтониан, характеризующий нелинейно-периодическую связь
между осцилляторами (
- гамильтониан, характеризующий нелинейно-периодическую связь
между осцилляторами (![]() - константа упругости цепочки,
- константа упругости цепочки, ![]() ),
), ![]() - гамильтониан,
- гамильтониан,
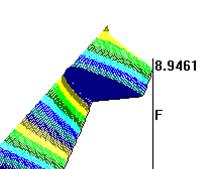 а)
б)
а)
б)
а)![]() б)
б) ![]()
а)x0=200 б)x0=250
Рис.3
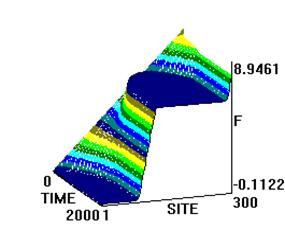 в)
в) 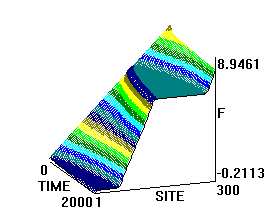 г)
г)
в)![]() г)
г) ![]()
в) x0=300 г) x0=350
Рис. 4
описывающий нелинейную связь между
“активной” и “замороженной” (![]() ) цепочками ДНК (
) цепочками ДНК (![]() - константа упругости водородных связей между
комплементарными основаниями, коэффициенты
- константа упругости водородных связей между
комплементарными основаниями, коэффициенты ![]() в уравнении (1) определяются в соответствии с правилом:
в уравнении (1) определяются в соответствии с правилом: ![]() в случае АТ и ТА пар,
в случае АТ и ТА пар, ![]() в случае ГЦ и ЦГ пар;
в случае ГЦ и ЦГ пар; ![]() - параметр, полученный ранее (см. выше) и определяемый
на основе модели синус-Гордона).
- параметр, полученный ранее (см. выше) и определяемый
на основе модели синус-Гордона).
При малых ![]() гамильтониан
гамильтониан![]() , что совпадает с соответствующей частью общего
гамильтониана, использованного ранее (см. выше). В этом случае уравнения
движения для
, что совпадает с соответствующей частью общего
гамильтониана, использованного ранее (см. выше). В этом случае уравнения
движения для ![]() , полученные из (1),
, полученные из (1),
имеют вид:
![]() (2)
(2)
где
произведена замена ![]() .
.
В случае ![]() в системе (2) можно перейти к безразмерному
дифференциальному уравнению синус-Гордона:
в системе (2) можно перейти к безразмерному
дифференциальному уравнению синус-Гордона:
![]() , (3)
, (3)
”непрерывный аналог” системы (2). Это уравнение имеет солитонные решения, в частности, односолитонное решение, или кинк, характеризующий динамику распространения дислокации в цепи.
В соответствии с (1) система нелинейных уравнений движения записывается следующим образом:
![]() (4)
(4)
Как видим, системы (2) и (4) существенно различаются. Отметим, однако, что проведенное численное моделирование динамики систем (2) и (4) показало следующее: если в качестве начальных условий для численного интегрирования (2) выбрать односолитонное решение его “непрерывного аналога” (3) - кинк (см. выше), то обнаруживается принципиальное сходство в характере решений.
Однако, при задании начальных условий в следующем виде:
 (5)
(5)
где ![]() -
”ступенчатая”
функция с высотой ступени
-
”ступенчатая”
функция с высотой ступени ![]() и
углом наклона уступа A, выявилось различие динамики данных систем
(срав. рис.1 и 2, 3). Более точно, системы (2) и (4) численно интегрировались
методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде
(7), в интервале
и
углом наклона уступа A, выявилось различие динамики данных систем
(срав. рис.1 и 2, 3). Более точно, системы (2) и (4) численно интегрировались
методом Рунге-Кутта четвертого порядка с начальными условиями, заданными в виде
(7), в интервале ![]() с шагом
с шагом ![]() .
Граничные условия - “квази-циклические”:
.
Граничные условия - “квази-циклические”:
![]()
![]() (поли-A-последовательность).
Параметр системы
(поли-A-последовательность).
Параметр системы ![]() . Варьировали параметр A
(угол наклона уступа функции
. Варьировали параметр A
(угол наклона уступа функции ![]() ).
).
Численное
интегрирование системы (2) показало, что образуются две уединенных волны,
движущихся справа налево по цепи с постоянной скоростью. Первая волна имеет
форму квазикинка, а вторая волна имеет форму квазибризера, причем скорость
первой волны превосходит таковую для второй. Обе волны за счет
“квазициклических” граничных условий, доходя до левого конца, появляются на
правом конце без изменения своей формы. Квазикинк, проходя по цепи маятников,
изменяет координату каждого маятника на угол ![]() (маятник
делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К
раз, он изменяет координату каждого маятника на угол
(маятник
делает полный оборот). Поэтому, проходя по замкнутой цепи маятников К
раз, он изменяет координату каждого маятника на угол ![]() Этим
объясняется “уступообразная” форма графиков. На рис. 2 представлены результаты
интегрирования системы (4) при тех же условиях. Из рисунка видно, что
образуются те же две уединенных волны - квазикинк и
квазибризер. Но принципиальное отличие от рассмотренного случая состоит в том,
что квазикинк в самом начале движется с отрицательным ускорением, так что в
результате его скорость оказывается меньше скорости квазибризера. Заметим, что
исследования проводились на однородной поли-A-последовательности; так что
изменение скорости квазикинка нельзя объяснить влиянием неоднородности цепочки.
Этот эффект объясняется нелинейным взаимодействием между ее мономерами.
Этим
объясняется “уступообразная” форма графиков. На рис. 2 представлены результаты
интегрирования системы (4) при тех же условиях. Из рисунка видно, что
образуются те же две уединенных волны - квазикинк и
квазибризер. Но принципиальное отличие от рассмотренного случая состоит в том,
что квазикинк в самом начале движется с отрицательным ускорением, так что в
результате его скорость оказывается меньше скорости квазибризера. Заметим, что
исследования проводились на однородной поли-A-последовательности; так что
изменение скорости квазикинка нельзя объяснить влиянием неоднородности цепочки.
Этот эффект объясняется нелинейным взаимодействием между ее мономерами.
Рис. 3 иллюстрирует результаты интегрирования системы (4) при тех же условиях за исключением того, что A=2. В данном случае реализуется только квазикинк и его отрицательное ускорение в начале движения таково, что в результате он движется в направлении, противоположном первоначальному. При интегрировании системы (2) в аналогичных условиях также образуется только квазикинк. Его скорость не меняется по сравнению со случаем Рис. 1.
Существенно, что при соответствующих условиях в системе типа ДНК или РНК могут возникнуть перевзбужденные ровибронные состояния. На квантовом языке это было бы адекватно перезаселению высоко лежащих квантовых уровней по сравнению с основным (реализации инверсной заселенности). В этом случае возникает заманчивая мысль, связанная с принципиальной возможностью создания биосолитонного лазера (БСЛ) на молекулах ДНК[4].
Однако, в
теории динамики биополимеров хорошо известно, что конформационные движения
реализуются по механизму ограниченной диффузии ввиду сильного влияния
диссипативных сил со стороны микроокружения. По этой причине решение проблемы
создания БСЛ на ДНК представляется весьма проблематичным, по крайней мере, для
подтверждения идеи необходимо выполнение условий: ![]() где
где ![]() и
и ![]() ‑ ширина и скорость солитона соответственно,
‑ ширина и скорость солитона соответственно, ![]() ‑ время диссипации. Положив
‑ время диссипации. Положив ![]()
![]() 5A и
5A и ![]() (скорость звука), имеем оценку
(скорость звука), имеем оценку ![]() . Отметим, что характерное время диссипации за счёт водных
гидродинамических сил
. Отметим, что характерное время диссипации за счёт водных
гидродинамических сил ![]() а время затухания, обусловливаемое процессами
внутри самой молекулы
а время затухания, обусловливаемое процессами
внутри самой молекулы ![]() (см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39.
С.949.; Чернавский и др. 1986. № 287. С. 21.).
(см., напр., Шайтан К.В. Биофизика. М.,1994. Т.39.
С.949.; Чернавский и др. 1986. № 287. С. 21.).
Существует также и другая сложность в отношении самосогласования биосолитонов и волны электромагнитного переизлучения. Напомним, что математическое моделирование в данном случае проводилось на монотонной поли-A ДНК и поэтому оставалось неясным влияет ли гетерогенная естественная последовательность ДНК на динамику солитонного возбуждения в молекуле. Чтобы проверить это, как и ранее, был взят С-район ДНК на 3’-конце вируса саркомы птиц в качестве полигона для запуска солитонов на разных участках полимера. На этот раз вычисляли производную от функции с тем, чтобы нагляднее показать движения солитонов.
Как и на Рис.1-3, на Рис.5-7 хорошо видны существенные модификации в поведении солитонов при изменениии параметра А. Особенно это заметно на рис.7, где солитонная волна идет, как и на Рис.5-6, вначале налево, а затем резко сворачивает направо. В этом видится определенный биологический смысл. Солитон как потенциальный “субъект чтения” ДНК должен “просматривать” протяженные контекстные зоны, а не застревать, синусоидально качаясь, на одних и тех же “словах” - локусх ДНК или РНК.
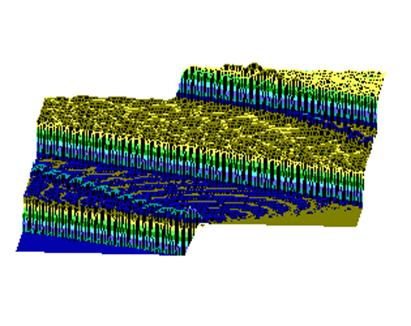 а)
а) б)
б)
Рис.5
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (2) при значении параметра A=1.
б) То же, вид сверху.
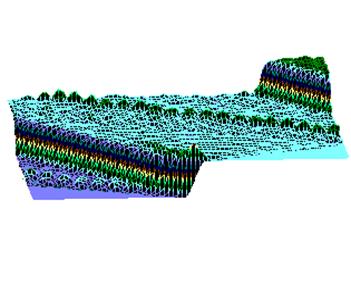 а)
а)
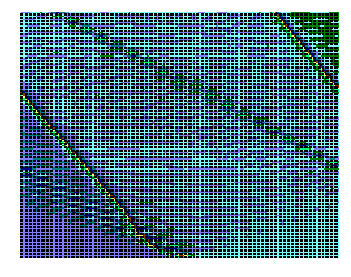 б)
б)
Рис.6
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=1.
б) То же, вид сверху.
 a)
a)
 б)
б)
Рис.7
а) Результаты численного моделирования динамики распространения возмущений в ДНК на основе системы (4) при значении параметра A=2.
б) То же, вид сверху.
Если же учитывать нелинейность ковалентных связей в сахарофо-фосфатном остове ДНК, то в поведении солитонов обнаруживаются дополнительные черты (рис. 8-10): сдвиг области инициации солитонной волны в ДНК-фрагменте из вируса саркомы птиц с 200-го до 500-го нуклеотида, то обнаруживаются дополнительные крутильные волны колебаний, расходящиеся в обе стороны от основной волны возмущения. Они, отталкиваясь от фиксированных концов ДНК (in vivo фиксаторами могут быть нуклеосомы), возвращаются к центральному возмущению и дополнительно модулируют его. Такие дополнительные волны могут играть роль “информаторов” о нуклеотидном составе и последовательности оснований в зондируемом участке ДНК или РНК, и эта информация может “запоминаться” на уровне явления возврата Ферми-Паста-Улама и использоваться хромосомным биокомпьютером для принятия соответствующих “решений”.
200-ый
Рис.8
Солитонное возбуждение ДНК, но с учётом нелинейности ковалентных связей в сахаро-фосфатном остове ДНК. Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 200-ый нуклеотид.
400-ый
Рис.9
То же, что на рис. 8, но центр возмущения - 400-ый нуклеотид.
500-ый
Рис.10
То же, что на рис. 9, но центр возмущения - 500-ый нуклеотид.
Существенная черта солитонных «сканирований» ДНК, особенно хорошо просматривающаяся на Рис. 8-10, - это наличие, кроме основной, дополнительных траекторий движения солитонов с богатой модуляцией. Такие дополнительные модулированные траектории солитонов, имеющие кинковую и бризерную структуру, могут нести дополнительные тонкие нюансы распределения волновой генетической информации вдоль цепей ДНК и РНК.
Антенная модель
Мы отмечали ранее [Гаряев, Маслов и др., 1996 (а); Гаряев, Маслов и др., 1996 (б)], что в роли, если не первичными, но одними из главных молекул, воспринимающих внешние электромагнитные поля как регуляторные, являются белки. И особенно металл содержащие. Функционирование ряда биологических макромолекул (в частности, ферментов) во многом определяется процессами, происходящими в активных центрах, окруженных биополимерными цепочками, имеющими знаковую топологию. Исходя из такого представления о структуре информационных биомакромолекул, естественно предположить, что их взаимодействие с физическими полями внешних по отношению к биосистеме и внутренних (организменных) излучений приводит к возбуждению дипольно-активных колебаний мономеров, формирующих указанную цепочку, а те, в свою очередь, индуцируют колебания в активном центре. Иными словами, такая система будет работать как своеобразная антенна. Эти возбужденные колебания способны привести к переходу биомакромолекулы в другое конформационное (топологическое, знаковое) состояние.
Подобная концепция в принципиальном плане адекватна целому ряду функционально высокозначимых биомакромолекул, например, хлорофилла, гемоглобина, миоглобина и т. д. Эти макромолекулы объединяются двумя структурными качествами: 1) в их геометрическом центре расположен ион (в случае хлорофилла - ион магния, в случае гемоглобина - ион железа); 2) около иона симметрично расположены 4 пиррольных кольца (псевдоплоская структура).
Другими типами биополимеров, соответствующих антенной модели, могут быть cравнительно простые циклы типа валиномицина (переносчик ионов калия) и сложные надмолекулярные структуры хромосом, ДНК которых содержит высокоорганизованные ассоциаты таких металлов, как магний, кальций, никель, кобальт, медь, железо, цинк и др. При этом роль их неясна и сводится исследователями, в основном, к нейтрализации ОН-групп остатков фосфорной кислоты полинуклеотида. Представляется, что функции металлов в ДНК и РНК существенно более широкие и реализуются по линии знакового и (или) энергетического взаимодействия с эндогенными и экзогенными по отношению к биосистеме физическими полями. То же относится и к белкам, не содержащим порфириновый центр, но специфическим образом связывающим металлы. Например, таковыми можно считать сайт-специфические белки с доменами типа “цинковых пальцев”, участвующими в регуляции генов, подчас очень далеко отстоящих от этих управляющих белков. Атомы металлов ДНК и белков могут резонансно взаимодействовать по электромагнитным каналам в рамках понятий антенной модели. Еще раз обозначим понятие антенной модели.
Внешняя энергия (в частности, связанная с резонансным взаимодействием крайне высокочастотных электромагнитных излучений с белками) поступает на периферию, т. е. на ансамбль субъединиц (не обязательно идентичных по структуре). В результате активной “беседы”, предопределенной биохимическими связями, между периферийными акцепторами (получившими закодированную энергию) и центром-ассоциатом (в данном случае ионом металла гемсодержащих белков), последний получает энергию (информацию), что и вызывает биологическое действие. Степень реакционной способности биомакромолекул существенно зависит от уровня возбуждения центральных субъединиц. Рассмотрим в деталях потенциальные механизмы волновых взаимодействий физических полей и активных центров информационных биомакромолекул в рамках предлагаемой нами антенной модели.
В качестве простейшей модели для иллюстрации антенного эффекта рассмотрим двумерную замкнутую (циклическую) цепочку мономеров. В центре цикла расположен активный центр, связанный с мономерами цепочки диполь-дипольным взаимодействием.
Обозначим
координатные смещения мономеров через ![]() , а смещение активного центра через
, а смещение активного центра через ![]() . Для потенциальной функции имеем:
. Для потенциальной функции имеем:
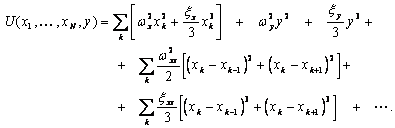 (1)
(1)
Первые два члена в (1) соответствуют колебаниям мономеров (второй член учитывает ангармонизм); последние два члена отвечают за связи между мономерами, Остальные члены отвечают за связи между мономерами и активным центром.
Уравнения движения запишем в виде:
![]() (2)
(2)
где ![]() - внешняя монохроматическая сила,
действующая только на мономеры,
- внешняя монохроматическая сила,
действующая только на мономеры, ![]() - коэффициент затухания, введенный феноменологически (простоты
ради принят одинаковым и для мономеров, и для активного центра).
- коэффициент затухания, введенный феноменологически (простоты
ради принят одинаковым и для мономеров, и для активного центра).
С учетом (1), система уравнений (2) приобретает вид:
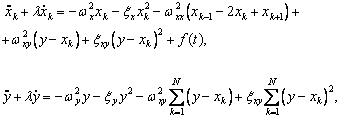 (3)
(3)
 (4)
(4)
Введем общую координату для ансамбля мономеров
![]() .
(5)
.
(5)
тогда система уравнений (4) в линейном приближении приобретает вид:
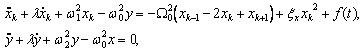 (6)
(6)
где:
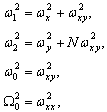
![]() - число мономеров.
- число мономеров.
С учетом (5)
имеем ![]() (7.1)
(7.1)
![]() (7.2)
(7.2)
Из (7.2)
следует ![]() (8)
(8)
Подстановка (8) в (7.1) дает
![]() . (9)
. (9)
Соответствующее
характеристическое уравнение имеет вид (после подстановки ![]() в однородное уравнение):
в однородное уравнение):
![]() (10)
(10)
Обозначив ![]() имеем
имеем
![]()
так что
![]() (11)
(11)
В дальнейшем предполагается выполнение неравенств:
![]() (12)
(12)
Первое условие соответствует случаю слабой связи между мономерами и активным центром, второе - малому затуханию мономерных осцилляторов.
Для собственных значений имеем
![]() , (13)
, (13)
где введены коллективные частоты:
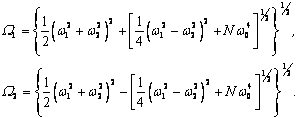 (14)
(14)
Нас
интересуют вынужденные колебания (внешняя сила ![]() ):
):
![]() . (15)
. (15)
Подстановка
(15) в (9) и приравнивание соответствующих коэффициентов при ![]() и
и ![]() дают систему алгебраических уравнений:
дают систему алгебраических уравнений:
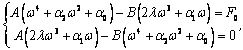
где:
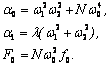
В результате
получаем ![]()
где 
После несложных, но громоздких преобразований для вынужденных колебаний активного центра получаем:
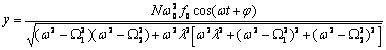 . (16)
. (16)
Из (16)
видно, что наибольшая амплитуда вынужденных колебаний активного центра достигается
в условиях коллективного резонанса: либо ![]() , либо
, либо ![]() .
.
В любом из этих случаев для амплитуды вынужденных колебаний имеем:
![]() (17)
(17)
Из (17) следует, что наибольший эффект резонансной раскачки активного центра достигается при большем числе периферийных субъединиц “антенны”, при более высоком значении коэффициента связи активного центра с мономерами, при наименьшем коэффициенте затухания и при наименьшем дисбалансе коллективных мод.
Нетрудно определить и “хореографию” (динамику вынужденных колебаний) отдельных мономерных единиц. В соответствии с (6) уравнение для k -го мономера запишем в виде:
![]() (18)
(18)
Вводя коллективные координаты
![]()
и применяя метод линейной алгебры, получаем для вынужденных колебаний мономеров:
![]() , (19)
, (19)
где:

![]() ‑ определяется из (16).
‑ определяется из (16).
Таким
образом, в рамках антенной модели наибольший эффект воздействия внешнего
монохроматического поля ![]() реализуется в условиях коллективного резонанса:
реализуется в условиях коллективного резонанса:
![]() .
.
Повторяя рассуждения раздела 2, можно сделать также следующие выводы:
1) При реализации амплитудной модуляции внешнего сигнала имеют место дополнительные возможности резонансного воздействия на биомакромолекулы на частотах:
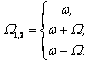
2) Учет
нелинейности при квадратичной связи для монохроматического сигнала привносит
дополнительный резонанс на второй гармонике ![]()
3) Учет нелинейности при амплитудной модуляции определяет еще ряд резонансных возможностей:

Таким образом, при действии резонансного электромагнитного поля на биомакромолекулы с активным центром, содержащим атомы металлов, существенную роль играют коллективные волновые эффекты. В этом случае свойства самого излучения предопределяют широкие возможности регуляторного влияния на динамику биомакромолекул в целом и, следовательно, на биопроцессы, в которых они принимают участие, тем самым прямо или косвенно реализуя управляющие и (или) дезорганизующие сигналы.
Литература
Albada P. van, Lagendijk A., 1985, Observation of Weak Localization of Light in a Random Medium, Phys. Rev. Lett. v.55, p. 2692-2695.
Bennet C.H., Brassard G., Crepeau C., Jossa R., Peres A., Wootters W.K., 1993, Teleporting and unknown quantum state via dual classical and Einstein-Podolsky-Rosen channels. Phys.Rev.Lett.,v.70, p.1895-1899.
Bouwmeester D., Pan Jian-Wei, Mattle K., Eibl M., Weinfurter H., Zeilinger A., 1997, Experimental quantum teleportation. Nature, v.390, p.575-579.
Einstein A., Podolsky B., Rosen N., 1935, Can quantum-mechanical description of physical reality be considered complete? Phys.Rev. v.47, p.777-780.
Fermi E., Pasta J., Ulam S., 1955, Studies of nonlinear problems. 1. Physics. Report. http://www.osti.gov/accomplishments/pdf/A80037041/A80037041.pdf
Gariaev P.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A., 1991, Hologrphic Associative Memory of Biological Systems, Proceedings SPIE - The International Society for Optical Engineering. Optical Memory and Neural Networks., v.1621, p.280- 291. USA.
Gariaev P.P., Birshtein B.I., Iarochenko A.M., Marcer P.J., Tertishny G.G., Leonova K.A., Kaempf U., 2001, The DNA-wave biocomputer. “CASYS” – International Journal of Computing Anticipatory Systems (ed. D.M.Dubois), Liege, Belgium, v.10, pp.290-310; http://www.rialian.com/rnboyd/dna-wave.doc
Gariaev P.P., 2003, Der wellengenetische Code. Tattva Viveka, №20, pp.68-73. (in German).
Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).
Lushnikov A.A., Maksimenko V.V.,.Simonov A.J, 1982, Electromagnetic Surface Modes in Small Metallic Particles, in Electromagnetic Surface Modes, ed. by A.D.Boardman, J. Wiley, Chichester, pp. 305-345.
Maksimenko V.V., 1999, Antoine’s Localization of Photon inside Fractal Cluster, Fractal in Engineering, Delft, Netherlands, p. 355-358.
Shcherbak V.I., 2003, Arithmetic inside the universal genetic code. BioSystems, v.70, pp.187–209.
Александров С.А. Некогерентный метод получения голограмм. // Оптика и спектроскопия. 1998, Т.85, № 6, С. 1029-1032.
Березин А.В., Курочкин Ю.А., Толкачев Е.А., 2003, Кватернионы в релятивистской физике. М.: УРСС.
Берсукер И.Б., 1976, Электронное строение и свойства координационных соединений. Ленинград. Химия.
Bischof M. Biophotonen. Das Licht in unseren Zellen. Printed in Germany, 1995, ISBN 3-86150-095-7, ZWEITAUSENDEINS, S. 522.
Благодатских В.И., Гаряев П.П., Леонова Е.А., Маслов М.Ю., Шайтан К.В., Щеглов В.А., 1996, О динамике возникновения дислокаций в молекуле ДНК. Краткие сообщения по физике. Физический Институт РАН, N3-4, с.9-14
Болтянский В.Г., Ефремович В.А, 1982, Наглядная топология, Москва, Наука, с. 84.
Борен К., Хафмен Д., 1986, Поглощение и рассеяние света малыми частицами. Москва, Мир, с. 77.
Бучаченко А.Л., 1979, Радиоизлучение и другие магнитные эффекты в химических реакциях. - М.: «Знание».
Будаговский А.В. Трансляция генетической информации посредством когерентных электромагнитных полей // Проблемы интенсификации современного садоводства, - Мичуринск, 1990, - С. 172-174.
Будаговский А.В., Евсеева Р.П. Экспериментальная модель дистанционной передачи морфогенетической информации в системе двух растительных тканей с разной потенцией к дифференцировке // Механизмы действия сверхмалых доз. Тез. Докл. 11 Межд. Симп. – М., 1995, - С. 124-125.
Будаговский А.В., Евсеева Р.П., Муратова С.А. Применение голограммы дифференцированного органа для индукции морфогенеза в культуре каллусных тканей плодовых растений // Биология растений in vitro, биотехнология и сохранение генофонда. Тез докл. Межд. Конф. – М., 1997. – С. 79.
Будаговский А.В., Туровцева Н.И., Будаговский И.В. Когерентные электромагнитные поля в дистанционном межклеточном взаимодействии // Биофизика. – 2001. – Т. 46, № 5 – С. 894-900.
Бурлаков А.Б., Бурлакова О.В., Голиченков В.А. Дистантные взаимодействия разновозрастных эмбрионов вьюна // ДАН. – 1999. – Т. 368. - № 4. – С. 562-564.
Будаговский А.В. Дистанционное межклеточное взаимодействие. НПЛЦ «ТЕХНИКА», М., 2004, С. 103.
Гаряев П.П., 1994, Волновой геном. М. Изд. Общественная польза. 279с.
Гаряев П.П., Маслов М.Ю., Решетняк С.А., Щеглов В.А., 1996 (а), Модель взаимодействия электромагнитного излучения с информационными биомакромолекулами., Краткие сообщения по физике. Физический Институт РАН, N1-2, с.60-63.
Гаряев П.П., Маслов М.Ю., Решетняк С.А., Щеглов В.А., 1996 (б), Взаимодействие электромагнитного излучения с информационными биомакромолекулами. "Антенная" модель. Краткие сообщения по физике. Физический Институт РАН, N1-2, с.54-59.
Гаряев П.П., Тертышный Г.Г., Готовский Ю.В., 1997, Трансформация света в радиоволны. III международная конференция «Теоретические и клинические аспекты применения адаптивной резонансной и мультирезонансной терапии». «ИМЕДИС». Москва. 18-20 апреля 1997г. с.303-313.
Гаряев П.П., 1997, Волновой генетический код. 107с. М. Издатцентр.
Гаряев П.П., Тертышный Г.Г., Приоритет на патент от 06.01.1999г. №99/01/Л (Федеральный институт промышленной собственности. Группа получающего ведомства РСТ. Старфилд, ЛТД).
Гаряев П.П., Тертышный Г.Г., Леонова Е.А., Максименко В.В., 2000, Волновой антивирусный иммунитет http://www.genetic-4.com/info/rus/s_imm.htm
Гаряев П.П., Кокая А.А., Мухина И.В., Леонова-Гаряева Е.А., Кокая Н.Г., 2007, Влияние модулированного биоструктурами электромагнитного излучения на течение аллоксанового сахарного диабета у крыс. Бюллетень Экспериментальной Биологии и Медицины, №2, с.155-158.
Денисюк Ю.Н. Об отображающих свойствах бегущих волн интенсивности при записи динамических объемных голограмм // ЖТФ, 1974, 44, №1, с. 131-136.
Кадомцев Б.Б., 1999, Динамика и информация, Москва, Редакция журнала “Успехи физических наук”, 400 с.
Киркин А.Ф. Нехимические дистантные взаимодействия между клетками в культуре // Биофизика. – 1981. Т. 26. – Вып. 5. – С. 839-843.
Mаксименко B.B.,. Крикунов В.А,. Лушников А.А, 1992, Сильная локализация света в плотноупакованных гранулированных средах, ЖЭТФ, т.102 , c.1571.
Мулдашев Э.Р., 2000, Комбинированная трансплантация глаза. Министерство здравоохранения, Российской Федерации, Всероссийский Центр Глазной и Пластической Хирургии. «Аллоплант». г. Уфа.
Налимов В.В., 1989, Спонтанность сознания. Вероятностная теория смыслов и смысловая архитектоника личности. Москва. "Прометей". 287с.
Мазур А.И., Грачев В.Н., 1985, Электрохимические индикаторы. М.: Радио и связь.
Молчанов А.А. Влияние световых межклеточных взаимодействий на интенсивность экструзии белка секреторными клетками молочной железы // Вестник ЛГУ. Биология. – 1985. - № 3. – Вып. 1. – С. 70-74.
Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Леонова Е.А., Мологин А.В., Гарбер М.Р., 2000(а), Генетические структуры как источник и приемник голографической информации. Датчики и Системы, №2, с.2-8.
Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Максименко В.В., Мологин А.В., Леонова Е.АМулдашев., Э.Р., 2000(б), Спектроскопия радиоволновых излучений локализованных фотонов: выход на квантово-нелокальные биоинформационные процессы. Датчики и Системы, №9 (18), с.2-13. http://www.wavegenetics.jino-net.ru/zip/Teleport.zip
Смелов М.В., 2001, Электромагнитные солитоны вакуума. Часть 4. Предварительные экспериментальные исследования, базирующиеся на модели электромагнитных солитонов. Физическая Мысль России. М.: МГУ. Т. 1. С. 38.
Cоломоник А., 1995, Семиотика и лингвистика. М., с. 345.
Тертышный Г.Г., Гетманов В.Г., Кузнецов П.А. Применение аппроксимационных алгоритмов в лазерном компьютерном виброметре // Измерительная техника, 1997, № 7, С. 34-37.
Тертышный Г.Г., Гетманов В.Г., Кузнецов П.А. Лазерный компьютерный виброметр. Тр. Межд. семинара «Вибродиагностика в промышленности » НПО «Спектр», М., 1998, С. 237-243.
Тертышный Г.Г, Гетманов В.Г., Дятлов А.В., Жиров М.В.,. Применение локальных и слайновых аппроксимаций для оценивания нестационарных параметров опто-электронных сигналов. ж. Автоматика и телемеханика, № 6, 2000, с. 29-35.
Чукова Ю. П., 2002, Эффекты слабых воздействий. – М.: Компания «Алес».