КЛЕТОЧНАЯ ОРГАНИЗАЦИЯ НЕРВНОЙ СИСТЕМЫ В.В. Жуков, Е.В. Пономарева
КЛЕТОЧНАЯ ОРГАНИЗАЦИЯ НЕРВНОЙ СИСТЕМЫ В.В. Жуков, Е.В. Пономарева
http://tom.imm.uran.ru/vos/1/0.htm
Калининград
1998
ВВЕДЕНИЕ
Нейроанатомия органически входит в состав нейробиологии - одной из наиболее бурно прогрессирующих областей человеческого знания. В анатомии этот прогресс коснулся прежде всего представлений о тонком строении нервной системы, о клеточном и субклеточном уровне ее организации. Как и многие другие биологические дисциплины, нейробиология - конкретная наука, требующая от изучающего ее определенных знаний анатомического субстрата.
Создавая это пособие, авторы постарались включить в него основные нейроанатомические понятия и термины, преследуя прежде всего цель - дать студентам конспект-справочник, облегчающий чтение нейробиологической литературы. Исходя из того, что эволюционное развитие нервной системы проходило во многом под влиянием возникновения и изменения сенсорных органов, мы поместили в пособие раздел, посвященный их строению. Кроме того, в пособие включено описание системы кровоснабжения центральной нервной системы, которое часто отсутствует в учебниках по нейробиологии. Основная часть содержания пособия касается анатомии нервной системы человека. Тем не менее студент найдет и краткое описание основных этапов филогенеза ее структуры. Иллюстративный материал пособия содержит расшифровку только упоминающихся в тексте структур. Ориентируясь на студентов-биологов, авторы сознательно избегали латинской номенклатуры. Отчетливо сознавая, что предлагаемое вниманию пособие не может быть единственным или главным источником учебного материала при изучении дисциплин нейробиологического цикла, авторы надеются, что их труд принесет реальную пользу всем, интересующимся различными аспектами строения нервной системы.
1. КЛЕТОЧНАЯ ОРГАНИЗАЦИЯ
НЕРВНОЙ СИСТЕМЫ
Нервная клетка
(нейрон)
Нервная клетка (нейрон) состоит из тела, от которого отходит один
или несколько отростков. Она обладает способностью проводить и передавать
электрические импульсы.
Сома, или тело, нейрона является центральным образованием, обеспечивающим рост дендритов и аксонов в эмбриогенезе, а также регенерацию аксона. У самых крупных нейронов диаметр сомы достигает 100 мкм и более, у самых мелких - около 5 мкм.
Дендритная зона - рецепторная мембрана, состоящая из сужающихся к концу цитоплазматических выростов (дендритов), с которыми образуются синаптические контакты других нейронов либо которые дифференцируются в структуру, трансформирующую воздействия внешней среды в электрическую активность.
Аксон - одиночный, нередко ветвящийся и удлиненный вырост цитоплазмы, структурно и функционально приспособленный для проведения нервных импульсов от дендритной зоны. У позвоночных животных он может иметь миелиновую оболочку, образованную клетками глии.
Телодендрии аксона - разветвленные и различно дифференцированные окончания аксонов, которым присуща мембранная и цитоплазматическая специализация, связанная с синаптической передачей или нейросекреторной активностью.
Строение
нейрона
Плазматическая мембрана окружает нервную клетку. Она состоит из белковых и липидных компонентов, находящихся в жидкокристаллическом состоянии (модель мозаичной мембраны): двуслойность мембраны создается липидами, образующими матрикс, в котрый частично или полностью погружены белковые комплексы. Плазматическая мембрана регулирует обмен веществ между клеткой и ее средой, а также служит структурной основой электрической активности.
Ядро отделено от цитоплазмы двумя мембранами, одна из которых примыкает к ядру, а другая к цитоплазме. Обе они местами сходятся, образуя поры в ядерной оболочке, служащие для транспорта веществ между ядром и цитоплазмой. Ядро контролирует дифференцировку нейрона в его конечную форму, которая может быть очень сложной и определяет характер межклеточных связей. В ядре нейрона обычно находится ядрышко.
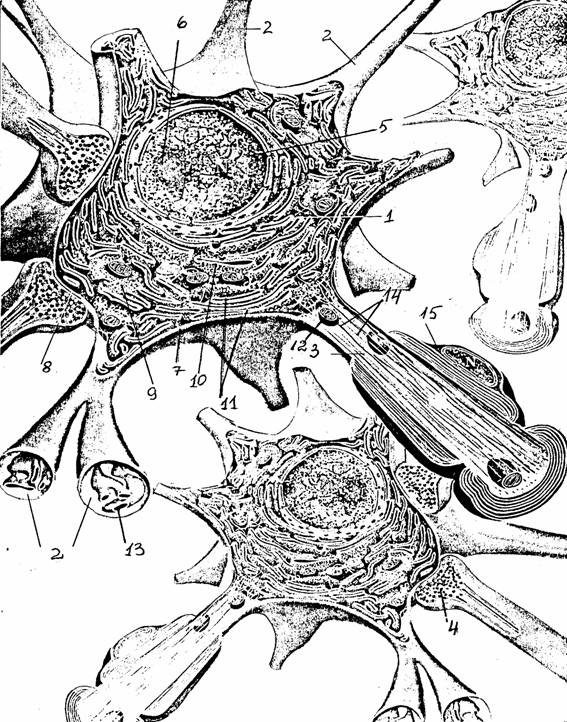
Рис. 1. Строение
нейрона (с
изменениями по [13]):
1 - тело
(сома), 2 - дендрит, 3 - аксон, 4 - аксонная терминаль, 5 - ядро,
6 -
ядрышко, 7 - плазматическая мембрана, 8 - синапс, 9 -
рибосомы,
10 -
шероховатый (гранулярный) эндоплазматический ретикулум,
11 -
субстанция Ниссля, 12 - митохондрии, 13 - агранулярный эндоплазматический ретикулум, 14 - микротрубочки и нейрофиламенты,
15 -
миелиновая оболочка, образованная шванновской
клеткой
Рибосомы производят
элементы молекулярного аппарата для большей части клеточных функций: ферменты,
белки-переносчики, рецепторы, трансдукторы,
сократительные и опорные элементы, белки мембран. Часть рибосом находится в
цитоплазме в свободном состоянии, другая часть прикрепляется к обширной
внутриклеточной мембранной системе, являющейся продолжением оболочки ядра и
расходящейся по всей соме в форме мембран, каналов, цистерн и пузырьков (шероховатый эндоплазматический ретикулум). В нейронах близ ядра образуется характерное
скопление шероховатого эндоплазматического ретикулума
(субстанция Ниссля), служащее местом интенсивного синтеза белка.
Аппарат Гольджи - система уплощенных мешочков, или цистерн - имеет внутреннюю, формирующую, сторону и наружную, выделяющую. От последней отпочковываются пузырьки, образующие секреторные гранулы. Функция аппарата Гольджи в клетках состоит в хранении, концентрировании и упаковке секреторных белков. В нейронах он представлен более мелкими скоплениями цистерн и его функция менее ясна.
Лизосомы - заключенные в мембрану структуры, не имеющие постоянной формы, - образуют внутреннюю пищеварительную систему. У взрослых особей в нейронах образуются и накапливаются липофусциновые гранулы, происходящие из лизосом. С ними связывают процессы старения, а также некоторые болезни.
Митохондрии имеют гладкую наружную и складчатую внутреннюю мембраны и являются местом синтеза аденозинтрифосфорной кислоты (АТФ) - основного источника энергии для клеточных процессов - в цикле окисления глюкозы (у позвоночных). Большинство нервных клеток лишено способности запасать гликоген (полимер глюкозы), что усиливает их зависимость в отношении энергии от содержания в крови кислорода и глюкозы.
Фибриллярные структуры: микротрубочки (диаметр 20-30 нм), нейрофиламенты (10 нм) и микрофиламенты (5 нм). Микротрубочки и нейрофиламенты участвуют во внутриклеточном транспорте различных веществ между телом клетки и отходящими отростками. Микрофиламенты изобилуют в растущих нервных отростках и, по-видимому, управляют движениями мембраны и текучестью подлежащей цитоплазмы.
Синапс - функциональное
соединение нейронов, посредством которого происходит передача электрических
сигналов между клетками. Щелевой
контакт обеспечивает электрический механизм связи между нейронами (электрический
синапс).
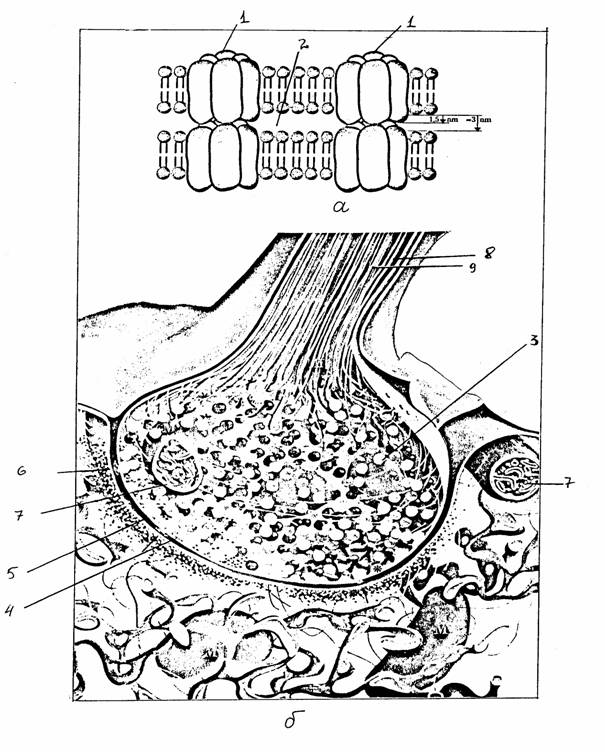
Рис. 2.
Строение синаптических
контактов:
а -
щелевого контакта, б - химического синапса (с изменениями по [13]):
1 - коннексон, состоящий из 6 субъединиц, 2 - внеклеточное
пространство,
3 - синаптическая везикула, 4 - пресинаптическая мембрана, 5 - синаптическая
щель, 6 -
постсинаптическая мембрана,7 - митохондрия, 8 -
микротрубочка,
9 - нейрофиламенты
Химический синапс отличается ориентацией мембран в направлении от нейрона к нейрону, что проявляется в неодинаковой степени уплотненности двух смежных мембран и наличием группы небольших везикул вблизи синаптической щели. Такая структура обеспечивает передачу сигнала путем экзоцитоза медиатора из везикул.
Синапсы также классифицируются в
зависимости от того, чем они образованы: аксо-соматические, аксо-дендритные, аксо-аксонные и дендро-дендритные.
Морфологические типы
нейронов
Униполярные клетки у беспозвоночных находятся в сенсорных узлах и в той и или иной степени связаны с сенсорными модальностями. У позвоночных подобные клетки имеют два отростка, которые сливаются вблизи тела клетки (псевдоуниполярные нейроны). Биполярные нейроны имеют один аксон и один дендрит и характерны для сенсорных органов зрительной, слуховой и обонятельной систем. Мультиполярные клетки имеют один аксон и несколько дендритов. Более подробная классификация исходит из особенностей формы их тела (веретенообразные, пирамидные), аксонной арборизации (корзинчатые), дендритного дерева, (звездчатые) и т.д. (всего до 60 различных вариантов).
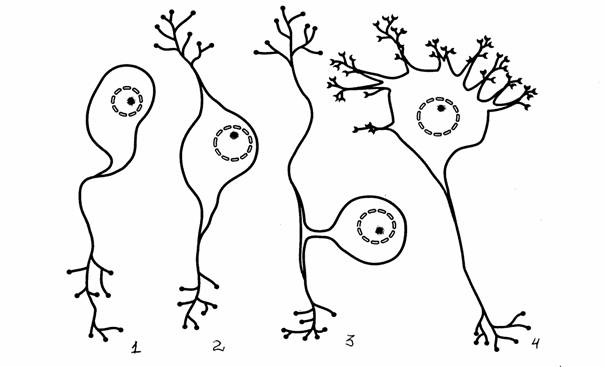
Рис. 3.
Морфологические типы нейронов [13]:
1 -
униполярный, 2 - биполярный, 3 - псевдоуниполярный, 4
- мультиполярный
Клетки
нейроглии
В некоторых отделах нервной системы клеток нейроглии почти в 10 раз больше, чем нервных. Астроциты обладают множеством отростков, в белом веществе мозга они носят название фиброзных (из-за наличия множества фибрилл в цитоплазме их тел и ветвей), а в сером веществе - протоплазматических. Они выполняют следующие функции: 1) служат опорой для нервных клеток; 2) обеспечивают репарацию нерва после повреждения; 3) изолируют и объединяют нервные волокна и окончания; 4) участвуют в метаболических процессах. Олигодендроциты имеют значительно меньше ветвей, они образуют миелиновые оболочки вокруг аксонов в ЦНС позвоночных. Оболочки периферических нервов образуются шванновскими клетками. Мелкие клетки микроглии рассеяны по всей нервной системе и фагоцитируют продукты распада.
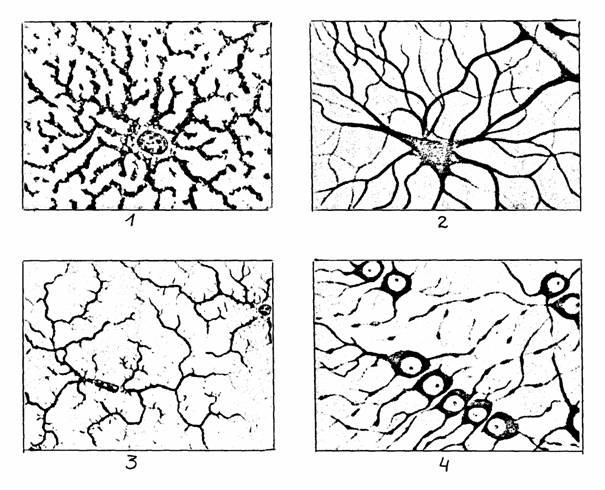
Рис. 4.
Клетки нейроглии (цит. по [12]):
1 -
протоплазматический астроцит, 2 - фиброзный астроцит,
3 - микроглия, 4 - олигодендроциты
2. СРАВНИТЕЛЬНАЯ
АНАТОМИЯ
НЕРВНОЙ СИСТЕМЫ
БЕСПОЗВОНОЧНЫХ
Тип стрекающих (Cnidaria)
Основа клеточной организации стрекающих - два слоя клеток (эктодерма и энтодерма), между которыми расположен слой неклеточного вещества мезоглии (диплобластические животные). В последнем располагаются нервные клетки, имеющие эктодермальное и энтодермальное происхождение. Отростки этих клеток, распространяясь на различные расстояния и образуя между собой синаптические контакты, образуют двумерную нервную сеть, проходящую по всему телу животного. Ее входами служат различные сенсорные клетки: сенсорные ямки (хеморецепция), глазки (точечные глаза), статоцисты (гравитационная чувствительность) или тактильные рецепторы. Отростки нервных клеток идут к сократительным эпителиальным клеткам, расположенным под колоколом медузы, что обеспечивает рефлексы, восстанавливающие положение тела в пространстве. Нервные клетки обладают спонтанной активностью, которая распространяется по нервной сети медленно и диффузно и обеспечивает плавательные движения. У некоторых видов медуз нервные клетки объединяются в небольшие нервные скопления (краевые тельца). Отсутствие выраженной центральной нервной системы (ЦНС) у стрекающих позволяет отнести ее нервную систему к диффузному типу структурной организации.
Тип плоские черви (Plathelminthes)
У плоских червей появляется третий зародышевый слой - мезодерма - признак характерный для всех других вышестоящих животных (триплобластические организмы). У примитивных плоских червей нервные клетки образуют нервную систему, мало отличающуюся от нервной системы стрекающих. У более продвинутых форм в головной части имеется скопление нервных клеток, знаменующее появление центральной нервной системы. От головного “мозга” отходят продольные нервные тяжи, отдающие в поперечном направлении нервы (ортогон). Хемо- и механорецепторы распределены по поверхности тела. На головном конце имеется группа вкусовых клеток и “глаза” - небольшие ямки с расположенными в них фоточувствительными клетками. При общей ограниченности форм поведения плоские черви демонстрируют простейшие ориентировочные рефлексы. У паразитических форм наблюдается частичная или полная редукция органов чувств и нервных клеток.
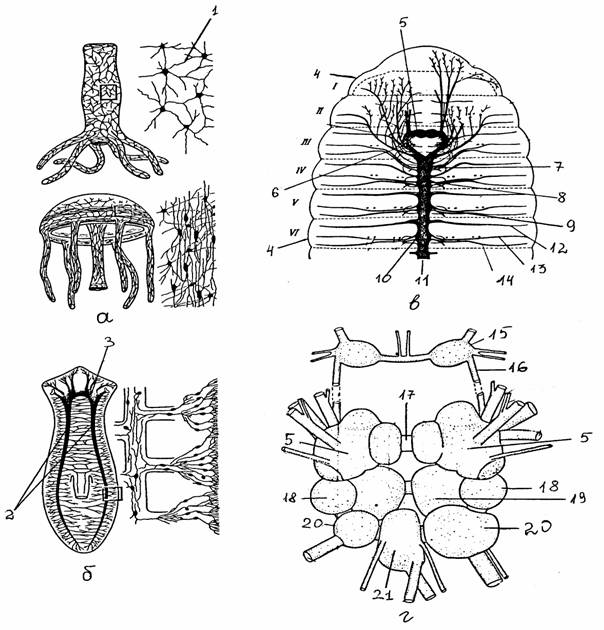
Рис. 5.
Строение нервной системы стрекающих,
плоских
червей, кольчатых червей, моллюсков:
а - нервная система гидры и
медузы [8]; б - нервная система планарии [8];
в - нервная система дождевого
червя - вид со спинной стороны (цит. по [2]);
г - ЦНС моллюска прудовика
[1]
1 -
диффузная сеть, 2 - продольные нервные тяжи, 3 - головной «мозг»
(церебральный
ганглий), 4 - сегменты тела
(I - YI), 5 - церебральный
ганглий,
6 -
окологлоточная коннектива, 7-10 - ганглии III,
IY, Y, YI сегментов
соответственно, 11 - брюшная нервная цепочка, 12, 13, 14 - передний,
средний, задний
сегментарные нервы, 15 -
буккальный ганглий, 16 - церебробуккальная коннектива,
17 - церебральная комиссура, 18 - плевральный ганглий, 19 -
педальный
ганглий, 20
- париетальный ганглий, 21 - висцеральный ганглий
Тип круглые черви (Nemathelminthes)
Триплобластические животные, имеющие первичную полость тела (щели между внутренними органами, непосредственно граничащие с окружающими тканями). Нервная система построена по типу ортогона. Основной группой типа является класс круглых червей (Nematoda). Центральная часть их нервной системы образована окологлоточным нервным кольцом, окружающим переднюю часть пищевода. С кольцом связана пара боковых головных ганглиев. От кольца вперед отходят 6 коротких нервных веточек, а назад направляются 6 нервных стволов. Среди последних два, проходящие по срединной спинной и брюшной линиям в соответствующих валиках гиподермы, наиболее мощные. Оба главных нервных ствола соединяются между собой многочисленными комиссурами, опоясывающими тело в виде полукольца попеременно с правой и левой сторон. Мускульный слой разбит валиками гиподермы на 4 продольные ленты. Спинной нервный ствол иннервирует мышцы обеих боковых спинных лент, брюшной - брюшных. Органы чувств нематод развиты слабо: имеются осязательные бугорки или щетинки, хеморецепторные органы, а также примитивные глаза - пигментные пятна или слабо развитый бокал.
Тип
кольчатые черви (Annelida)
Триплобластические целомические животные со сквозным кишечником и сегментированным телом. Нервные клетки сгруппированы в ганглии, образующие ЦНС. Ганглии головной части тела образуют мозг. В каждом метамере (сегменте тела) ганглии расположены попарно и билатерально, соединяясь комиссурами через среднюю линию. Соединение между метамерами осуществляется с помощью продольных тяжей, или коннектив, а с периферией - с помощью нервов. Ганглии, соединенные коннективами, образуют брюшную нервную цепочку. Нервные клетки расположены на наружной поверхности ганглиев и имеют по одному толстому отростку, ответвления которого идут в нейропиль и по коннективам и комиссурам направляются к другим ганглиям или в составе нервов идут на периферию. У некоторых червей имеется система гигантских нервных волокон, идущих через всю брюшную нервную цепочку. Такие гигантские аксоны встречаются также у головоногих моллюсков (отряд десятиногие), у членистоногих. У кольчатых червей есть рецепторы, обеспечивающие чувство равновесия, хеморецепторы, а также фоторецепторы, сосредоточенные у некоторых форм в камерных глазах.
Тип
моллюски (Mollusca)
Нервная система моллюсков может быть различной степени сложности и состоит из ганглиев, наиболее крупные из которых сгруппированы вокруг пищевода. Пары ганглиев соединяются между собой комиссурами, а с другими ганглиями - коннективами. Из органов чувств имеются статоцист и осфрадий (орган химического чувства), у многих хорошо развиты глаза. Нервную систему некоторых брюхоногих моллюсков легко изолировать, она содержит необычно крупные клетки, что делает ее излюбленным объектом нейробиологов. Центральная нервная система головоногих моллюсков достигает большой степени развития. Расположенные вокруг пищевода ганглии сильно увеличины и, сливаясь, образуют настоящий мозг. Очень высокого уровня развития достигают глаза. Зрительные ганглии превращаются в сложные зрительные доли - самые крупные отделы мозга. Нейроны этих долей дифференцируются на ряд форм, отличающихся от униполярных нейронов, характерных для беспозвоночных. Гигантский аксон нервной системы кальмара стал классическим экспериментальным объектом нейрофизиологиии.
Тип
членистоногие (Arthropoda)
К типу членистоногих относятся животные, имеющие состоящий из хитиновых пластинок-склертов наружный скелет и членистые конечности. Большая часть видов относится к насекомым, другие главные группы - паукообразные и ракообразные. Нервная система ракообразных содержит надглоточный (церебральный) и подглоточный ганглии, образующие головной мозг, и брюшную нервную цепочку. Есть сердечный ганглий, управляющий сокращениями сердца, и система гигантских нервных волокон. Главными среди рецепторных органов являются расположенные на подвижных стебельках сложные глаза, несущие хеморецепторы антенны и статоцисты. Имеется несколько типов механорецепторов.
У насекомых церебральные ганглии сливаются
и образуют несколько отделов: протоцеребрум,
получающий входы от глаз; дейтоцеребрум,
получающий вход от антенн; и тритоцеребрум, иннервирующий переднюю
часть
пищеварительного канала и область головы. Хорошо выделяются зрительные доли
протоцеребрума, что связано с высоким уровнем развития
сложных глаз. В грудном отделе расположены три крупных ганглия: про- мезо- и метаторакальный. Число брюшных ганглиев
варьирует (максимально до 8). Расположенные около аорты кардиальные тела
соединены с мозгом посредством нервного ствола. С ними связаны прилежащие тела.
И те и другие являются нейроэндокринными органами.
3.
СРАВНИТЕЛЬНАЯ АНАТОМИЯ
НЕРВНОЙ СИСТЕМЫ
ПОЗВОНОЧНЫХ
Общий план строения
Центральная нервная система (ЦНС)
построена из клеток и волокон, которые развились из дорсально расположенной нервной трубки. Периферическая нервная система - нервные
волокна, соединяющие ЦНС и тело, а также группы клеток, которые лежат за
пределами ЦНС и называются ганглиями. ЦНС делится на две основные части: спинной мозг, лежащий внутри
позвоночного столба, и головной мозг,
находящийся внутри черепной коробки и состоящий из 5 отделов. Спинномозговые нервы образуются
дорсальными и вентральными корешками спиного мозга.
Черепномозговые нервы (у высших позвоночных их 12 пар)
начинаются в полости черепа, а их ядра (кроме ядер I и II нервов) лежат в стволе. Вегетативная нервная система образует
систему моторной иннервации внутренних органов. Нейроэндокринная система осуществляет
связь между нервной системой и телом с помощью гормонов, ее основная часть - гипофиз.
Эволюция мозга
позвоночных
Эволюция сложных форм поведения позвоночных связана прежде всего с увеличением сложности ствола мозга, и особенно конечного мозга. Мозг примитивного позвоночного состоит из заднего, среднего и переднего отделов. В филогенезе над задним мозгом развился мозжечок, в результате чего задний мозг разделился на мост и продолговатый мозг. В среднем мозге тектум изменился для более сложной переработки зрительной и слуховой информации. В переднем мозге произошло интенсивное развитие покрывающего слоя (плаща), который вместе с базальными ганглиями образовал полушария большого мозга. У птиц и млекопитающих такое развитие большого мозга связано с поступлением зрительной, слуховой и соматосенсорной информации, а также с усложнением ее переработки и осуществлением сложных движений. Дифференцировка промежуточного мозга способствует выполнению двух важных функций: переключения информации на ее пути между полушариями и остальными отделами головного мозга и управления гипофизом, который в свою очередь управляет эндокринной системой организма. При всей сложности картины филогенетического развития мозга основной является тенденция к увеличению размера и сложности конечного мозга. Наиболее сильно меняются полушария большого мозга и мозжечок, что проявляется в увеличении числа извилин, приводящем к увеличению площади коры мозга.
4.
ФУНКЦИОНАЛЬНЫЕ ОТДЕЛЫ МОЗГА ЧЕЛОВЕКА
Спинной мозг
Спинной мозг находится внутри костного канала, образованного позвоночником, и удерживается в постоянном положении при помощи связок, идущих от мягкой и паутинной оболочек мозга к внутренней поверхности твердой оболочки. Последняя прикрепляется к позвонкам воронкообразными выростами в области межпозвоночных отверстий, через которые выходят корешки. Пространства между твердой, паутинной оболочками и мозгом (к которому плотно прилегает мягкая оболочка) заполнены спинномозговой жидкостью (СМЖ), которая также заполняет и центральный спинномозговой канал.
Характерная особенность организации
спинного мозга - наличие правильной периодичности в выходе задних (дорсальных) и передних (вентральных) спинномозговых корешков,
содержащих соответственно афферентные
(центростремительные) и эфферентные
(центробежные) волокна. На дорсальном корешке находится спинномозговой
ганглий, содержащий афферентные нервные клетки, отростки которых входят в
спинной мозг. Каждая пара корешков соответствует одному из позвонков и покидает
позвоночный канал через отверстия между ними. После выхода из костного канала
корешки сливаются и оба типа волокон смешиваются в нервах. Поэтому спинной мозг
разделен на ряд сегментов (шейные,
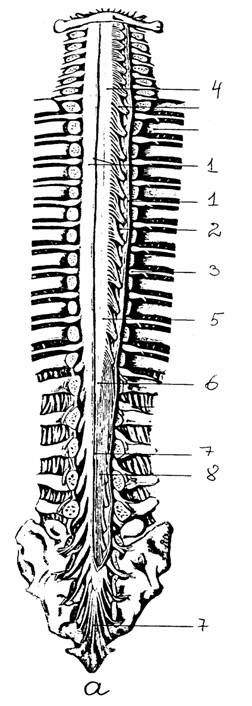
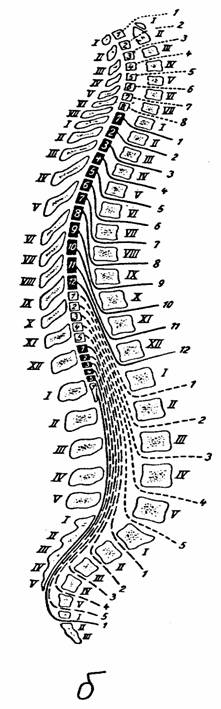
Рис. 7.
Спинной мозг в позвоночном канале [14]:
а - внешний
вид спинного мозга (твердая оболочка частично удалена):
1 - твердая
оболочка и ее край, 2 - зубчатая связка, 3 -
спинномозговой
ганглий, 4
- шейное утолщение, 5 - поясничное утолщение,
6 -
мозговой конус, 7 - концевая нить, 8 - конский хвост;
б - схема
расположения сегментов спинного мозга по отношению
к
позвоночному столбу: римские цифры - позвонки,
арабские
цифры - сегменты спинного мозга и спинномозговые
нервы
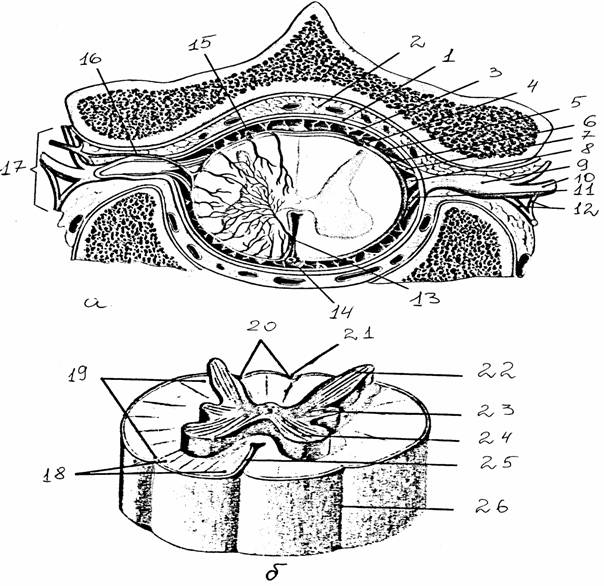
Рис. 8.
Строение спинного мозга [14]:
а -
поперечный срез через позвоночный канал на уровне 2-го грудного
позвонка;
б - белое и
серое вещество спинного мозга
1 - твердая
оболочка, 2 - эпидуральное пространство, 3 - субдуральное
пространство, 4 - паутинная
оболочка, 5 - субарахноидальное пространство,
6 - мягкая
оболочка, 7 - задний корешок, 8 - зубчатая связка, 9 -
спинномозговой ганглий, 10 - спинной нерв, 11 - передний корешок, 12 -
соединительные
ветви, 13 -
бороздчатокомиссуральная артерия, 14 - передняя
спинномозговая
артерия, 15
- задняя спинномозговая артерия, 16 - спинномозговая ветвь
межреберной
артерии, 17 - межпозвоночное отверстие, 18-20 - передний,
боковой,
задний канатики, 21 - задняя срединная борозда, 22-24 -
задний,
боковой,
передний столбы серого вещества, образующие на поперечном
сечении
рога (задние, боковые, передние - соответственно), 25 -
передняя
срединная щель, 26 - передняя боковая борозда
грудные, поясничные, крестцовые), число которых равно количеству позвонков. На третьем месяце внутриутробного развития плода спинной мозг занимает всю длину позвоночного канала. В последующие месяцы позвоночник растет в длину быстрее, чем спинной мозг, который у взрослых людей достигает 1-го поясничного позвонка. Каудальный конец спинного мозга имеет коническую форму (мозговой конус). Корешки нижних сегментов идут единым пучком, покрытым мозговой оболочкой (конский хвост). Отходящая от мозгового конуса тонкая фиброзная нить (концевая нить) прикрепляется к надкостнице крестца и фиксирует положение дистального конца спинного мозга. В поясничной и шейной области корешки образуют короткие смешанные нервы, формирующие поясничное и плечевое сплетения.
В спинном мозге сосредоточены нервные
клетки, аксоны которых дают начало нервам, идущим к поперечнополосатым мышцам
тела. Эти и другие нейроны образуют серое
вещество спинного мозга, расположенное вокруг его центрального канала. В нем
различают передние и задние рога, а также разделяющую их промежуточную часть. Задние рога
содержат нейроны, которые передают сенсорные импульсы к лежащим выше центрам или
расположенным вентральнее двигательным клеткам,
замыкая рефлекторные дуги. В передних рогах находятся самые крупные нейроны,
иннервирующие мышцы (двигательные
нейроны, или мотонейроны). В промежуточной зоне находятся клетки
серого вещества, связывающие его чувствительный и двигательный отделы. С 1-го
грудного сегмента спинного мозга до 3-го поясничного в боковых отделах серого
вещества расположены боковые столбы,
состоящие из двигательных нейронов симпатической части автономной нервной
системы. Расположение нейронов в сером веществе неравномерно: определенные их
типы собираются в группы (столбы, или
ядра).
Серое вещество спинного мозга окружено слоем белого вещества, состоящего из восходящих и нисходящих миелинизированных волокон. Белое вещество разделяется выростами серого на каждой стороне на три канатика: передний (вентральный), задний (дорсальный) и боковой (латеральный). Восходящие пути передают сенсорную информацию к рефлекторным подкорковым центрам и коре головного мозга, нисходящие проводят информацию к двигательным клеткам передних рогов. Системы коротких волокон (проприоспинальные пути) связывают между собой разные сегментарные уровни спинного мозга.
Проводящие пути
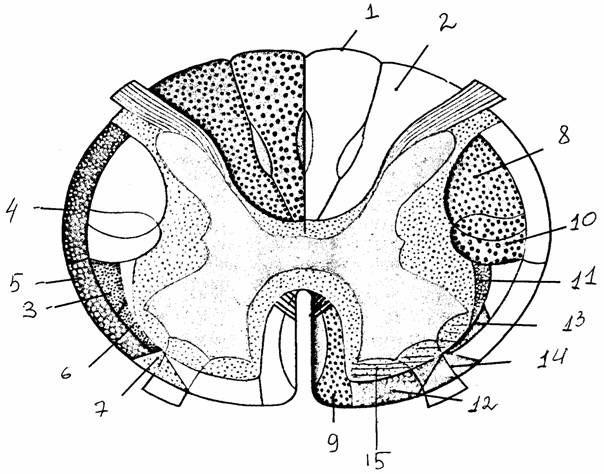
Рис. 9.
Проводящие пути спинного мозга
[14]:
восходящие
пути: 1 -
тонкий пучок, 2 - клиновидный пучок,
3 -
спинно-таламический путь, 4 - спинно-мозжечковый задний
путь,
5 -
спинно-мозжечковый передний путь, 6 - спинно-тектальный путь,
7 - спинно-оливарный путь;
нисходящие
пути: 8 - кортико-спинальный латеральный (пирамидный)
путь,
9 - кортико-спинальный передний (пирамидный) путь, 10 - рубро-спинальный
путь, 11,
12 - вестибуло-спинальные (латеральный и передний)
пути, 13 - текто-спинальный путь, 14 - оливо-спинальный путь, 15 - ретикуло-спинальный путь.
Восходящие пути
1. Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) проходят в дорсальных канатиках и образованы аксонами афферентных неронов спинномозговых ганглиев. Нейроны 2-го порядка находятся в продолговатом мозге в ядрах Голля (медиальные) и Бурдаха (латеральные). Аксоны нейронов этих ядер в стволе мозга совершают перекрест (медиальная петля). Нейроны 3-го порядка находятся в ядрах промежуточного мозга и направляют аксоны в кору больших полушарий. По этим путям в ЦНС поступает сенсорная информация главным образом от проприоцепторов мышц и сухожилий, а также от тактильных рецепторов кожи.
2. Спинно-таламический путь обеспечивает передачу в основном болевой и температурной чувствительности. Нейроны 1-го порядка располагаются в спинномозговых ганглиях, а 2-го порядка - в основании дорсальных рогов серого вещества спинного мозга. Аксоны последних переходят на противоположную сторону и по белому веществу латерального или вентрального канатика проходят без перерыва до промежуточного мозга. Расположенные в нем нейроны 3-го порядка посылают аксоны в кору больших полушарий.
3. Спинно-мозжечковые пути - дорсальный (пучок Флексига) и вентральный (пучок Говерса) - занимают латеральные канатики. Нейроны 1-го порядка также находятся в спинальных ганглиях. Аксоны нейронов 2-го порядка, расположенные в сером веществе спинного мозга, идут по латеральным канатикам той же (путь Флексига) или противоположной (путь Говерса) стороны к мозжечку. В стволе мозга волокна пути Говерса снова перекрещиваются и поступают в кору мозжечка той же стороны. Оба пути обеспечивают передачу сигналов от проприоцепторов мышц и сухожилий, а также висцерорецепторов.
Таким образом, кора больших полушарий связана с афферентными волокнами противоположной стороны тела, а кора мозжечка связана преимущественно с нейронными структурами одноименной стороны.
Нисходящие пути
1. Кортико-спинальный (пирамидный) путь образован аксонами пирамидных клеток V слоя преимущественно двигательных областей коры больших полушарий. Эти аксоны проходят без перерыва через ствол мозга. Большая часть их в области продолговатого мозга переходит на противоположную сторону (перекрещенный путь) и проходит в вентральном канатике до соответствующих сегментов спинного мозга. Небольшая часть волокон не претерпевает перекреста в продолговатом мозге, однако и они переходят на противоположную сторону в сегментах спинного. В обоих случаях двигательная кора больших полушарий оказывается напрямую связанной с нейронами противоположной стороны спинного мозга. Этот путь филогенетически один из самых молодых в ЦНС и развит только у млекопитающих. Он связывает пирамидные клетки двигательной коры, как правило, с интернейронами (у приматов также и с мотонейронами).
2. Рубро-спинальный путь образован аксонами нейронов красного ядра среднего мозга. Эти аксоны перекрещиваются непосредственно под ядром и по латеральному канатику спускаются до различных сегментов спинного мозга, устанавливая в его сером веществе связи с интернейронами. Это более древняя, чем пирамидный путь, система контроля двигательной функции и развита не только у млекопитающих, но и у рептилий и птиц.
3. Вестибуло-спинальный путь образован нейронами расположенных в продолговатом мозгу ядер Дейтерса. Их аксоны проходят в вентральном канатике и заканчиваются на интер- и мотонейронах промежуточной зоны серого вещества спинного мозга.
4. Ретикуло-спинальный путь образован аксонами ретикулярной
формации заднего мозга. Основная часть этих волокон заканчивается на интернейронах медиальной части вентрального рога.
Задний мозг (ромбовидный
мозг)
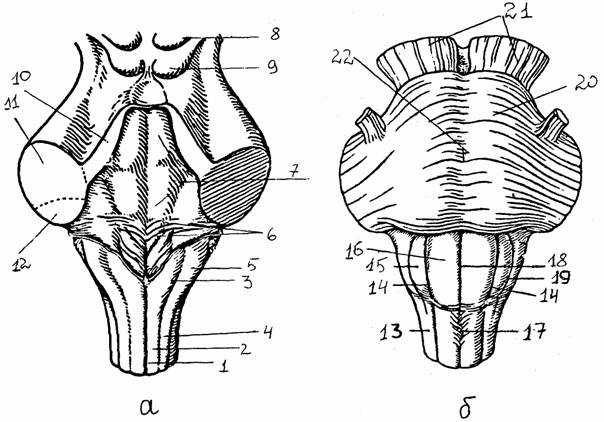
Рис. 10.
Ствол мозга [6]:
а - задняя
поверхность (мозжечок удален): 1 - задняя срединная
борозда,
2 - тонкий
пучок, 3 - бугорок тонкого ядра, 4 - клиновидный пучок,
5 - бугорок
клиновидного ядра, 6 - мозговые полоски, 7 - ромбовидная
ямка,
дно
IY желудочка, 8, 9 - верхние и
нижние бугорки четверохолмия,
10-12 -
верхняя, средняя, нижняя мозжечковые ножки;
б -
передняя поверхность: 13 - боковой канатик, 14 - передне-латеральная
борозда, 15
- олива, 16 - пирамида, 17 - перекрест пирамид, 18 -
передняя
срединная
щель, 19 - позади-оливная борозда, 20 - мост, 21 - ножки
мозга,
22 -
базилярная борозда
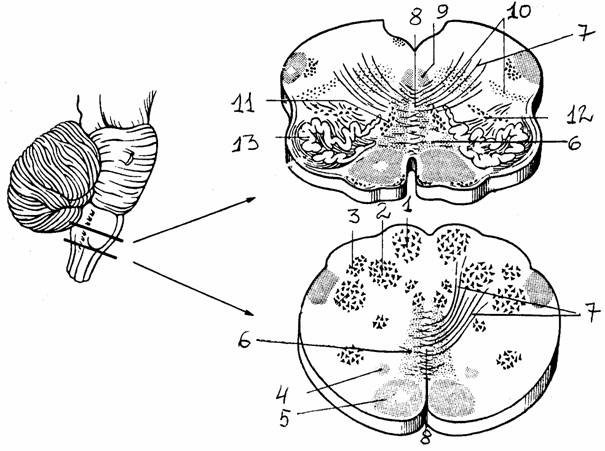
Рис. 11.
Поперечный разрез продолговатого мозга (на двух уровнях) [6]:
1 - тонкое
ядро, 2 - клиновидное ядро, 3 - добавочное клиновидное
ядро,
4 - текто-спинальный путь, 5 - пирамидный пучок, 6 - медиальная
петля,
7 -
внутренние дугообразные волокна, начинающиеся от тонкого и
клиновидного ядер, формируют медиальную петлю, 8 - перекрест
медиальных петель,
9 -
медиальный продолговатый пучок, 10 - ретикулярная формация,
11,
12 -
медиальное и заднее добавочные оливные ядра, 13 - нижнее оливное
ядро
Задний мозг включает в
себя продолговатый мозг и варолиев мост и сохраняет черты сегментарного
строения, что проявляется в расположении ядер V - XII пар черепно-мозговых нервов. Характерной
особенностью является наличие мощной массы нервных элементов, которые не
получают непосредственно афферентных волокон из периферических источников и не
отдают двигательных волокон, - ретикулярной формации. Эта надсегментарная структура занимает всю центральную часть
заднего мозга и доступна для вмешательства со стороны дна IV желудочка. Другие надсегментарные структуры - ядра Голля и Бурдаха, а также расположенные в толще мозга оливы.
Средний мозг
Средний мозг состоит из трех основных частей: крыша, покрышка и ножки мозга. Крыша образована парами верхних и нижних бугров четверохолмия. Между крышей и расположенной вентральнее покрышкой проходит сильвиев водопровод. В покрышке находятся ядра III и IV пар черепномозговых нервов, а также добавочное ядро III нерва (парасимпатическое ядро Эдингера - Вестфаля), парные красные ядра и несколько ядерных скоплений меньшего размера, относящихся к ретикулярной формации. Ножки мозга содержат главным образом нисходящие двигательные волокна, начинающиеся в коре мозга и идущие к ядрам черепномозговых нервов и двигательным ядрам спинного мозга. Непосредственно под красными ядрами находится группа клеток, составляющих черное вещество, анатомически входящее в состав ножек мозга.
Мозжечок
Основные части мозжечка - два полушария и непарный червь. Со стволовой частью мозжечок соединяется тремя парами ножек: верхними - со средним мозгом, средними - с варолиевым мостом, нижними - с задним мозгом. Его поверхность разделяется глубокими бороздами на доли и извилины. По Ларселу, поверхность мозжечка делят на отделы в зависимости от поступающих афферентных путей и филогенетического возраста. Филогенетически более древний архицеребеллум занимает наиболее каудальную часть (флоккуло-нодулярная доля). Палеоцеребеллум включает в себя переднюю часть червя, пирамиду, язычок и парафлоккулярный отдел. Масса мозжечка состоит из покрывающей его поверхность коры и расположенного глубже белого вещества с ядрами.
Кора состоит из трех слоев.
1. Поверхностный молекулярный состоит из параллельных
волокон и разветвлений дендритов нижележащих клеток. В нижней части расположены
тела корзинчатых клеток, аксоны которых оплетают клетки Пуркинье, а также
звездчатые клетки.
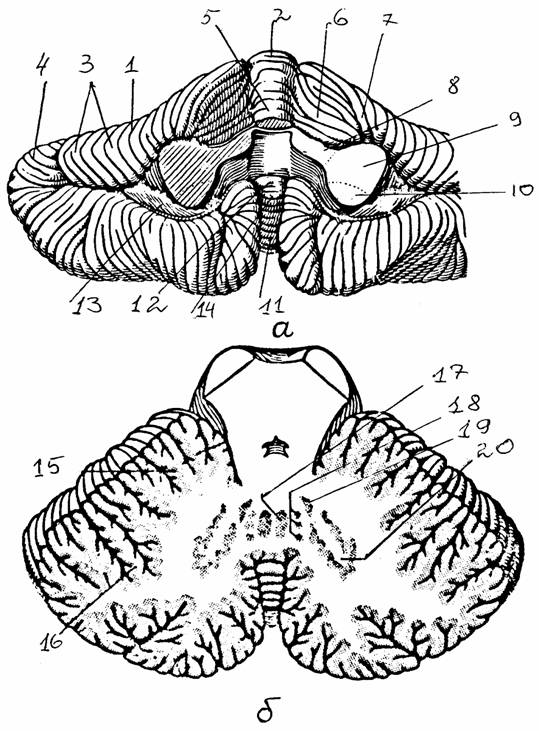
Рис. 13.
Строение мозжечка: а - вид снизу, б - горизонтальный разрез [6]:
1 -
полушарие мозжечка, 2 - вершина, 3 - листки мозжечка, 4 -
борозда,
5 - язычок,
6 - крыло центральной дольки, 7 - четырехугольная долька
(передняя
часть), 8-10 - верхняя, средняя и нижняя мозжечковые
ножки,
11 -
пирамида, 12 - узелок, 13 - клочок, 14 - язычок червя, 15 - кора
мозжечка,
16 - древо
жизни мозжечка, 17 - ядро шатра, 18 - шаровидное ядро,
19 - пробковидное ядро, 20 - зубчатое ядро
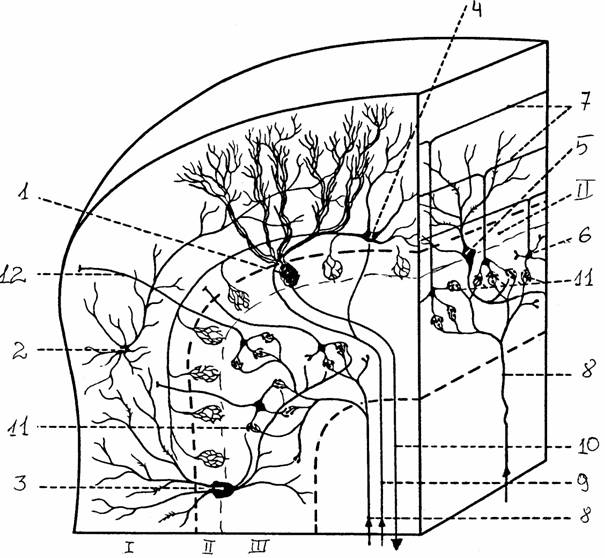
Рис. 14.
Кора мозжечка и ее нейронный состав (с изменениями, по
[15]):
I
- молекулярный
слой, II - ганглиозный слой, Ш - гранулярный
слой;
1 - клетка
Пуркинье, 2 - звездчатая клетка, 3 - клетка Гольджи,
4 - корзинчатая клетка, 5 - клетка Гольджи, 6 - клетка-зерно, 7 -
параллельные
волокна, 8
- мшистые волокна, 9 - лазающие волокна, 10 - аксон клетки
Пуркинье, 11 - гломерулы мозжечка, 12 - аксон зернистой
клетки
2. Ганглиозный слой - тела клеток Пуркинье. Эти клетки имеют обильно разветвленное дендритное дерево, которое направляется к поверхности мозжечка, и аксон, идущий в белое вещество. Аксоны клеток Пуркинье - единственные выход из коры мозжечка, они заканчиваются на нейронах ядер мозжечка и вестибулярных ядер.
3. Гранулярный слой содержит большое число тел клеток-зерен. Их аксоны поднимаются вертикально в молекулярный слой и там Т-образно ветвятся, образуя параллельные волокна, которые формируют синапсы на дендритах других клеток. В этом же слое находятся клетки Гольджи, аксоны которых подходят к клеткам-зернам.
Афферентный вход мозжечка образован тремя системами волокон. Во-первых, идущие из олив продолговатого мозга лазающие (лиановидные) волокна, заканчивающиеся синапсами на дендритах клеток Пуркинье. Во-вторых, мшистые (моховидные) волокна идут от ядер моста и заканчиваются на клетках-зернах. В-третьих, адренэргические волокна из голубого пятна, лежащего на границе заднего и среднего мозга.
Подкорковые парные ядра мозжечка: ядро шатра, шаровидное, пробковидное и зубчатое. Аксоны нейронов этих ядер идут к стволовым структурам: от ядер шатра - к вестибулярному ядру Дейтерса и ретикулярной формации, от шаровидных и пробковидных - к красным ядрам, от зубчатых - к таламусу и далее к коре больших полушарий.
Афферентные связи мозжечка различны в его отделах: архицеребеллум получает связи по вестибуло-мозжечковому пути, палеоцеребеллум - по спинно-мозжечковому пути, а неоцеребеллум - также по спинно-мозжечковым путям, по корково-мостовым путям, главным образом от сенсо-моторной коры больших полушарий.
Промежуточный
мозг
Клеточные скопления промежуточного мозга локализуются вокруг третьего желудочка, образуя его боковые, верхние и нижние стенки. Они могут быть поделены на 5 отделов: зрительный бугор (таламус), забугорная область (метаталамус), подбугорная область (гипоталамус), надбугорная область (эпиталамус), субталамическая область (субталамус).
В средней части обе половины таламуса соединяются между собой вместе перемычкой - промежуточной массой. Анатомически ядра таламуса делят на группы: передние, интраламинарные, срединные, боковые и задние. Ядра, дающие четко очерченные проекции в сенсорную кору больших полушарий, называют специфическими, или проекционными. В них заканчиваются длинные аксоны нейронов восходящих афферентных систем (кроме обонятельного пути). Неспецифические ядра связаны с ретикулярной формацией и дают в коре полушарий диффузные проекции. Выделяют также ассоциативные ядра, получающие сигналы от специфических, и моторные ядра, включенные в систему регуляции движений.
К метаталамусу относятся латеральные (ЛКТ) и медиальные (МКТ) коленчатые тела, являющиеся проекционными ядрами.
В состав эпиталамуса входит шишковидная железа (эпифиз) и ядра уздечки.
К субталамусу относится ядерное образование с тем же названием.
В гипоталамус входят: перекрест зрительных нервов,
заканчивающаяся гипофизом воронка, серый бугор и наиболее
каудально расположенные сосцевидные
тела.
|
Рис.
16. Ядра гипоталамуса (цит. по [15]): 1 -
передняя комиссура, 2 - конечная пластинка, 3 - преоптическая область, 4 -
вентромедиальное ядро, 5 - супраоптическое
ядро, 6 -
зрительный перекрест, 7 - зрительный нерв, 8 - гипоталамическая борозда, 9
- паравентрикулярное ядро, 10 - дорсомедиальное
ядро, 11 - заднее ядро, 12 -
сосцевидное тело, 13 - латеральные ядра серого
бугра, 14 -
ядра воронки, 15 - воронка, 16 -
нейрогипофиз, 17 - промежуточная доля, 18 -
передняя доля |
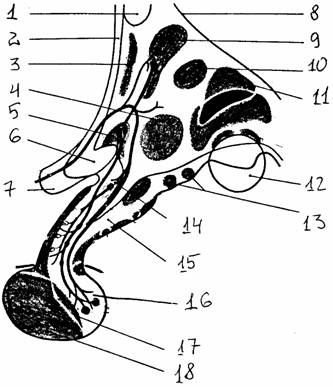
В этих образованиях расположено большое количество ядер, которые можно разделить на несколько групп: преоптическую, переднюю, среднюю, наружную и заднюю. Наиболее хорошо выделяются паравентрикулярное и супраоптическое ядра (передняя группа), заднее гипоталамическое и мамиллярное ядра (задняя группа). Среди сложных и запутанных связей гипоталамуса можно выделить короткий, но очень мощный пучок волокон (до 100 000 аксонов), заканчивающийся в задней доле гипофиза. По этим волокнам происходит не только распространение нервных волокон, но и ток нейросекрета, вырабатываемого клетками супраоптического и паравентрикулярных ядер. Гипофиз, являющийся важнейшей железой внутренней секреции, состоит из трех долей: передней (аденогипофиз), промежуточной и задней (нейрогипофиз) - из которых только последняя является производным нервной системы.
Таблица
1
Черепномозговые нервы и
их функции
|
Номер
пары |
Наименование
нерва |
Функция |
|
I |
Обонятельный |
Сенсорный
вход от обонятельного эпителия |
|
II |
Зрительный |
Сенсорный
вход от ганглиозных клеток
сетчатки |
|
III |
Глазодвигательный |
Моторный
выход к четырем из шести наружных мышц глазного
яблока |
|
IV |
Блоковый |
Моторный
выход к верхней косой мышце глазного яблока |
|
V |
Тройничный |
Основной
сенсорный вход от лица. Моторный выход к жевательным
мышцам |
|
VI |
Отводящий |
Моторный
выход к наружной прямой мышце глазного яблока |
|
VII |
Лицевой |
Основной
моторный выход к мышцам лица. Сенсорный вход от некоторых вкусовых
рецепторов |
|
VIII |
Слуховой |
Сенсорный
вход от внутреннего уха и вестибулярного
органа |
|
IX |
Языкоглоточный |
Сенсорный
вход от рецепторов (в том числе вкусовых) языка и
глотки |
|
X |
Блуждающий |
Главный
парасимпатический моторный выход к мышцам многих внутренних органов:
сердца, желудка, кишечника и др. Моторный выход к мышцам глотки. Сенсорный
вход от некоторых вкусовых рецепторов |
|
XI |
Добавочный |
Моторный
выход к грудино-ключично-сосковой и трапециевидной
мышцам |
|
XII |
Подъязычный |
Моторный
выход к мышцам языка |
Конечный мозг
Конечный мозг состоит из покрытых корой двух полушарий и базальных (подкорковых) ядер. У
взрослого человека поверхность полушарий делится на лобную, теменную, затылочную и
височную доли. Каждый из этих
отделов разделяется бороздами на
меньшие по размерам извилины: верхние,
средние и нижние височные и лобные, прецентральную
и постцентральную и т.д. Латеральная (сильвиева) борозда отделяет височную долю от теменной и
лобной, границу между которыми образует
центральная (роландова) борозда. Отдельно
расположен островок, лежащий в
глубине латеральной борозды на боковой поверхности полушария и прикрытый лобной,
височной и теменной долями. Выделяют также лимбическую долю, в которую входят части лобной,
теменной и височной долей: подмозолистая область, поясная извилина, перешеек поясной
извилины, парагиппокампальная извилина, крючок гиппокампа, миндалина и диагональная связка Брока.
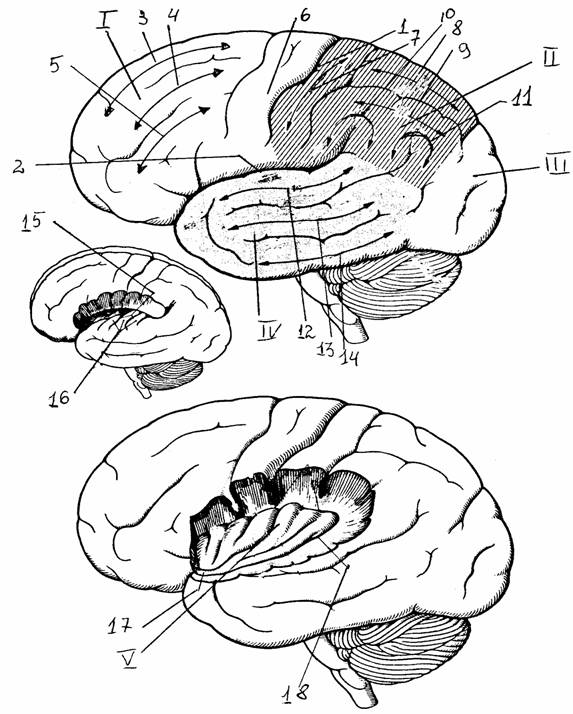
Рис. 17.
Латеральная поверхность полушарий [6]:
I - Y - лобная, теменная,
затылочная, височная и островковая
доли;
1 -
центральная борозда, 2 - латеральная борозда, 3, 4, 5 - верхняя,
средняя,
нижняя
лобные извилины, 6, 7 - прецентральная и
постцентральная извилины,
8, 9 -
верхняя, нижняя теменные дольки, 10 - надкраевая
извилина, 11 - угловая
извилина,
12, 13, 14 - верхняя, средняя, нижняя височные извилины,
15 -
поперечные височные борозды, 16 - поперечные височные
извилины
(Гешля), 17 - порог островка, 18 - круговая борозда
островка
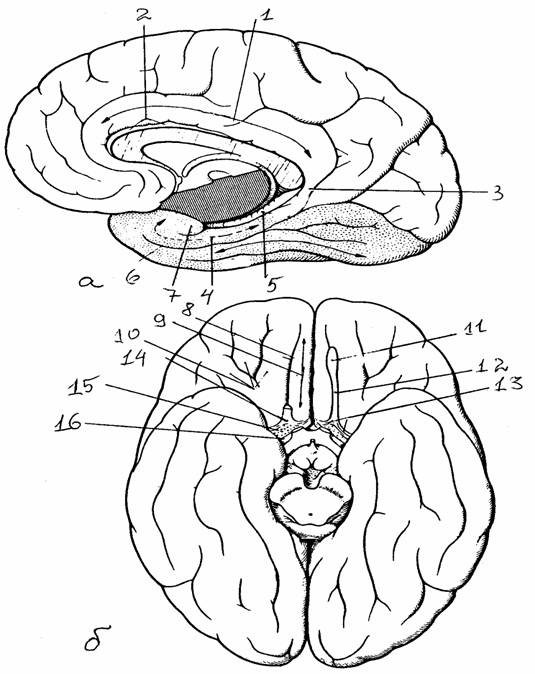
Рис. 18.
Полушария большого мозга:
а -
медиальная поверхность, б - основание полушарий
[6]:
1 - поясная
извилина, 2 - борозда мозолистого тела, 3 - перешеек
поясной
извилины, 4
- парагиппокампальная извилина, 5 - зубчатая
извилина,
6 - носовая
борозда, 7 - крючок, 8 - обонятельная борозда, 9 - прямая
извилина,
10 -
глазничные извилины, 11 - обонятельная луковица, 12 -
обонятельный
тракт, 13 -
медиальная и латеральная обонятельные полоски, 14 -
обонятельный
треугольник, 15 - переднее
продырявленное вещество, 16 - диагональная
извилина
Брока, в глубине которой проходит компактный пучок
волокон-
диагональная связка Брока
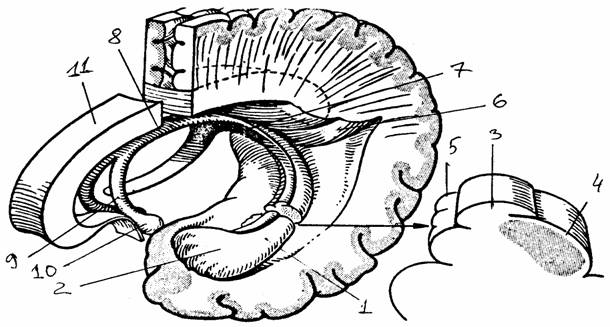
Рис. 19.
Гиппокамп левого полушария [6]:
1 - гиппокамп, 2 - ножка, 3 - бахромка, 4 - лоток, 5 - зубчатая
извилина,
6 - птичья
шпора, 7 - ножка свода, 8 - тело свода, 9 -столб,
10 -
сосцевидное тело, 11 - мозолистое тело
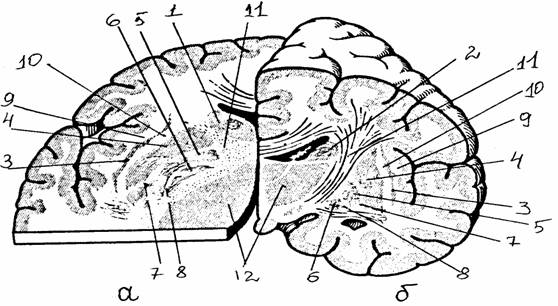
Рис. 20.
Базальные ядра на горизонтальном (а)
и
фронтальном (б) разрезах полушарий головного мозга [6]:
1, 2 - головка и тело
хвостатого ядра, 3 - ограда, 4 - скорлупа,
5, 6 -
латеральный и медиальный сегменты бледного шара, 7,8 -
наружная
и
медиальная мозговые пластинки, 9-11 - самая наружная,
наружная
и
внутренняя капсулы, 12 - таламус
Части коры млекопитающих содержат следы филогенетического развития. Палеокортекс: обонятельные луковицы (в которые непосредственно поступают волокна от обонятельного эпителия), обонятельные тракты (расположены на нижней поверхности лобных долей), обонятельные бугорки (вторичные обонятельные центры) и некоторые другие близлежащие структуры.
Архекортекс: поясная извилина, извилина морского коня (извилина гиппокампа), миндалевидное тело (скопление серого вещества под корой височной доли).
Остальные части коры относят к неокортексу, который у млекопитающих занимает доминирующее положение, оттесняя старую кору вперед и внутрь.
Комплекс базальных ядер (полосатое тело) расположен в толще конечного мозга и состоит из хвостатого ядра, бледного шара, скорлупы и ограды. Полоска белого вещества (внутренняя капсула), проходящая между таламусом и хвостатым ядром с одной стороны и бледным шаром - с другой. Наружная капсула отделяет полосатое тело от ограды.
Основной структурной особенностью коры является правильная ориентация клеточных элементов и волокон, которые идут в направлении перпендикулярном поверхности, либо параллельно ей. Обычно близкие по размеру клетки группируются на одинаковой глубине, образуя клеточные слои.
Типичной для млекопитающих является шестислойная кора: 1) верхний молекулярный (плексиморфный) слой содержит мелкие нервные клетки и апикальные волокна пирамидных клеток нижележащих слоев; 2) наружный зернистый слой содержит пирамидные клетки небольшого размера; 3) слой средних пирамид; 4) внутренний зернистый слой состоит из большого количества короткоаксонных нейронов; 5) внутренний пирамидный слой содержит особенно большие пирамидные нейроны; 6) полиморфный слой. Однако в различных участках коры имеются определенные различия в нейронной организации, что лежит в основе создания карт цитоархитектонического строения коры. Например, на карте Бродмана выделено 11 областей, включающих 52 поля.
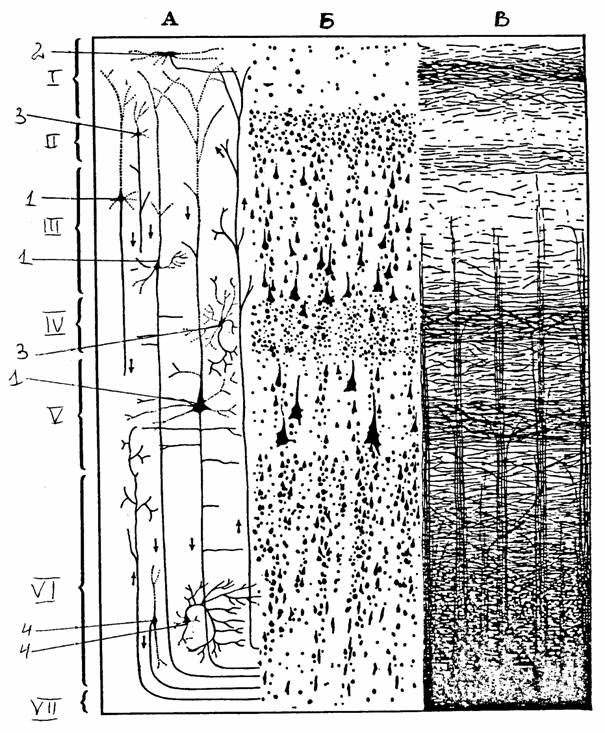
Рис. 21.
Строение коры больших полушарий [15]:
А - слои
коры, Б - цитоархитектоника, В - миелоархитектоника;
слои:
I
- молекулярный,
II - наружный
зернистый, III
- внешний
слой
пирамидных клеток (средних пирамид), IY
- внутренний
зернистый,
Y
- внутренний
пирамидный, YI
- полиморфный,
YII - белое
вещество;
1 -
пирамидные клетки разных размеров, 2 - клетка Кахаля,
3 -
звездчатые клетки, 4 - непирамидные клетки различного
типа
5. АНАТОМИЯ ВЕГЕТАТИВНОЙ
НЕРВНОЙ СИСТЕМЫ
Строение симпатической
системы
Центральная часть представлена ядрами, расположенными в грудных и верхних поясничных сегментах спинного мозга (латеральная часть промежуточной области серого вещества, образующая боковые рога). Аксоны нейронов (группа В, по Эрлангеру и Гассеру) этих ядер выходят из спинного мозга в составе вентральных корешков. После выхода из позвоночного канала эти аксоны идут в составе смешанного нерва и, отделяясь от него, в составе отдельных веточек (белых соединительных ветвей) подходят к ганглиям парных симпатических стволов, которые тянутся вдоль позвоночника. В этих ганглиях преганглионарные аксоны, как правило, образуют синаптические окончания на ганглионарных нейронах. Их постганглионарные аксоны идут дальше на периферию и иннервируют эффекторные органы: гладкую мускулатуру внутренних органов, железистые клетки, поперечнополосатую мускулатуру сердца. Постганглионарные волокна очень тонкие и, как правило, очень слабо миелинизированы (серые соединительные веточки). Количество ганглиев пограничного ствола превышает количество белых соединительных ветвей: они расположены на уровне не только грудных и верхних поясничных, но и шейных и нижних поясничных сегментов. В этих случаях ганглии не имеют собственных белых соединительных ветвей, а соединены с другими ганглиями продольными комиссурами. Эфферентные волокна проходят к ним из грудного отдела по продольным комиссурам через промежуточные ганглии, не прерываясь. В средних грудных сегментах значительная часть аксонов клеток бокового рога проходит через симпатический ствол без перерыва, образуя мощный чревный нерв. Он подходит к лежащим в брюшной полости ганглиям солнечного сплетения, где его волокна заканчиваются синаптическими окончаниями. Часть симпатических преганглионарных волокон заканчивается в более мелких ганглиях, расположенных на брыжейке и в полости таза.
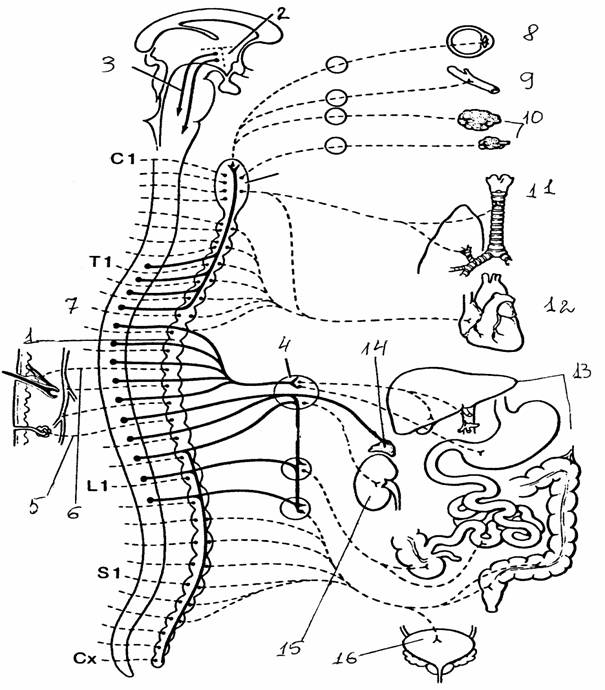
Рис. 23.
Органы, иннервируемые симпатической системой (цит. по
[9]):
Т1,
L1
- грудной,
поясничный отделы спинного мозга;
1 -
симпатический ствол, 2 - гипоталамус, 3 - нисходящие вегетативные
пути,
4 - чревный
ганглий, 5 - потовые железы, 6 - мускулатура волосяного
фолликула,
7 - периферические кровеносные сосуды, 8 - гладкая
мускулатура
глаза, 9 -
сосуды головы, 10 - железы, связанные с глазом, с носовой и
ротовой
полостями,
11 - дыхательная система, 12 - система кровообращения,
13 -
пищеварительная система, 14 - надпочечник, 15 - почка,
16 -
мочевой пузырь и органы размножения
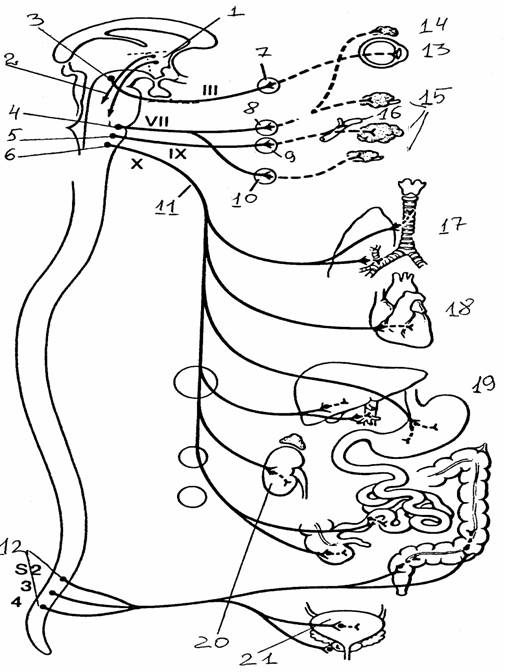
Рис. 24.
Органы, иннервируемые парасимпатической системой (цит.
по [9]):
1 -
гипоталамус, 2 - нисходящие вегетативные пути, 3 - добавочное
ядро
(ядро Эдингера-Вестфаля)
III нерва, 4 -
верхнее слюноотделительное
ядро
YII
нерва, 5 -
нижнее слюноотделительное ядро IX нерва, 6 - дорсальное ядро
X
нерва, 7 -
ресничный ганглий, 8 - крылонебный ганглий, 9 -
ушной
ганглий, 10
- поднижнечелюстной ганглий, 11 - блуждающий (Х)
нерв,
12 -
крестцовый отдел, 13 - гладкая мускулатура глаза, 14 - слезная
железа,
15 -
слюнные железы, 16 - сосуды головы, 17 - дыхательная
система,
18 -
система кровообращения, 19 пищеварительная система,
20 - почка,
21 - мочевой пузырь, органы размножения.
Наличие синаптического перерыва на периферии - морфологическая особенность вегетативной нервной системы, позволяющая четко дифференцировать ее эфферентный путь от соответствующего соматического. В симпатической системе есть только одно исключение - волокна, проходящие через симпатический ствол без перерыва и заканчивающиеся на клетках мозгового слоя надпочечников (хромаффинные клетки). Эти клетки имеют нейрогенное происхождение, и эмбриологически их можно рассматривать как видоизмененные ганглионарные нейроны.
Строение парасимпатической
системы
Центральные структуры парасимпатического
отдела расположены в среднем, заднем, продолговатом мозгу и крестцовом
отделе спинного мозга. Среднемозговая часть представлена добавочным ядром, заднемозговая - верхним слюноотделительным ядром. В
продолговатом мозге находятся два парасимпатических ядра - нижнее
слюноотделительное и дорсальное ядра. Часть преганглионарных волокон покидает ствол мозга в составе III,
VII и IX черепно-мозговых нервов и заканчивается на нейронах ресничного, ушного, крылонебного и челюстного узлов. Клетки преганглионарных нейронов дорсального ядра посылают свои
волокна в составе X пары нервов к органам шеи, грудной и брюшной полости.
Крестцовый отдел представлен центрами трех крестцовых сегментов спинного мозга
(S2 - S4). Аксоны
клеток этих центров выходят в составе вентрального корешка, присоединяются к
различным нервным стволам и направляются к периферическим ганглиям. Эти ганглии
располагаются непосредственно в иннервируемых органах или вблизи них: ободочная,
сигмовидная и прямая кишка, мочевой пузырь, мочеиспускательный канал, половые
органы.
6. ОБОЛОЧКИ МОЗГА
Самая наружная - твердая мозговая оболочка, образованная волокнистой соединительной тканью. В полости черепа она срастается с надкостницей, выстилающей поверхность костей черепа. Это сращение отсутствует там, где проходят крупные вены, отводящие кровь от мозга и заключенные между твердой оболочкой и надкостницей. В позвоночном канале твердая оболочка отделена от надкостницы скоплением жира и эпидуральным венозным сплетением. В полости черепа, в тех местах, где располагаются крупные венозные синусы, внутренний листок твердой оболочки отходит от стенки черепа. Эти листки идут несколько дальше заключенных в них венозных синусов и образуют плотные соединительнотканные пластинки, разделяющие некоторые отделы мозга.
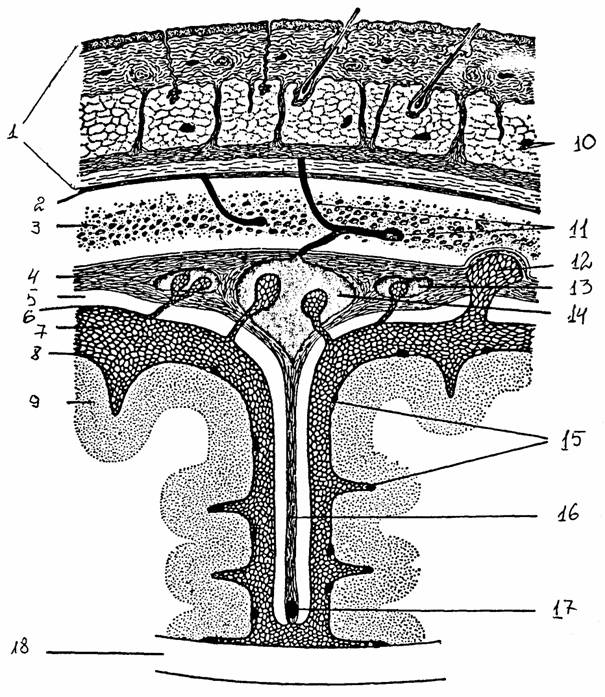
Рис. 25.
Оболочки головного мозга на фронтальном разрезе [15]:
1 - кожа, 2
- надкостница, 3 - кость черепа, 4 - твердая оболочка,
5 - субдуральное пространство, 6 - паутинная оболочка, 7 - подпаутинное
пространство, 8 - мягкая
оболочка, 9 - кора мозга, 10 - сосуды кожи,
11 - диплоические вены, 12 - грануляции паутинной оболочки, 13 -
лакуна
синуса
твердой оболочки, 14 - верхний сагиттальный синус, 15 -
сосуды
мягкой
оболочки, 16 - серп большого мозга, 17 - нижний
сагиттальный
синус, 18 -
мозолистое тело
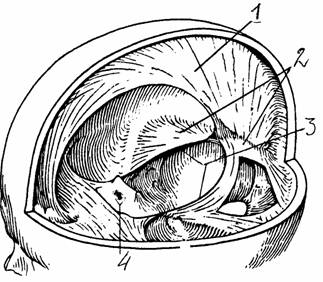
|
Рис.
26. Серп мозга и палатка (намет) мозжечка
[6]: 1 -
серп большого мозга, 2 -
намет мозжечка, 3 -
вырезка намета, 4 -
диафрагма седла |
Одна из них - серп большого мозга - расположена между двумя полушариями. Другая - палатка мозжечка - проходит горизонтально, отделяя мозжечок от большого мозга. Еще одна складка, диафрагма седла, изолирует гипофиз, лежащий на турецком седле.
Паутинная оболочка, образованная
эластичной волокнистой тканью, лежит под твердой оболочкой. Она отделена от
твердой промежутком - субдуральным пространством, которое в норме не
существует, так как паутинная оболочка плотно прилежит к твердой (“потенциальное
пространство”). Паутинная оболочка соединена с лежащей глубже мягкой оболочкой посредством тонких
перемычек. Эти две оболочки отделены друг от друга подпаутинным пространством, заполненным спинномозговой жидкостью. В некоторых
отделах подпаутинное пространство расширяется, образуя
цистерны.
Цистерны находятся над каждой бороздой или щелью поверхности мозга. Самая крупная из них - большая цистерна - находится между задне-нижней поверхностью мозжечка и продолговатым мозгом. Через твердую мозговую оболочку в интрадуральные венозные синусы проникают небольшие особые выросты паутинной оболочки, покрытые “шапочкой” из эпителиальных клеток. Это грануляции паутинной оболочки, через которые часть спинномозговой жидкости, заполняющей субарахноидальное пространство, может поступать в кровяное русло.
Мягкая мозговая оболочка плотно соединена с поверхностью головного и спинного мозга и связана с поверхностным слоем астроцитов. В ней находятся тонкие сплетения капилляров, которые служат для всасывания части спинномозговой жидкости.
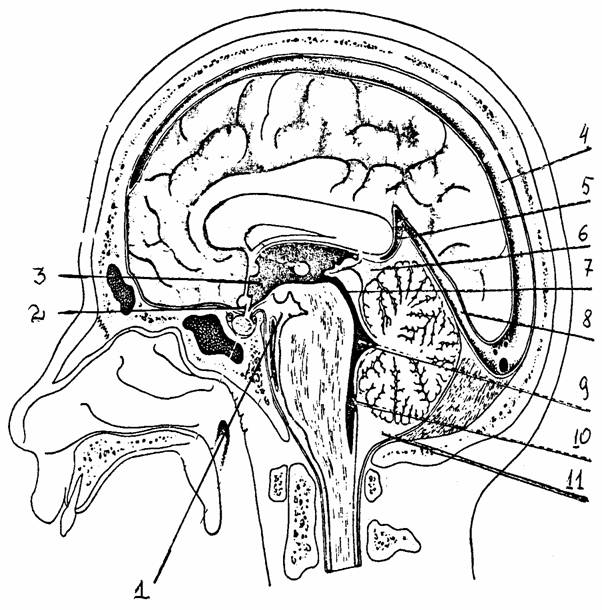
Рис. 27.
Цистерны подпаутинного пространства (цит. по [15]):
1
- межножковая
цистерна, 2 - цистерна перекреста, 3 - III
желудочек
мозга,
4 - верхний
сагиттальный синус, 5 - большая мозговая (Галена) вена,
6 -
охватывающая цистерна, 7 - водопровод мозга, 8 - прямой
синус,
9 -
IY желудочек мозга, 10 -
срединное отверстие IY
желудочка,
11 -
мозжечково-мозговая (большая) цистерна
7. СИСТЕМА ЖЕЛУДОЧКОВ
МОЗГА
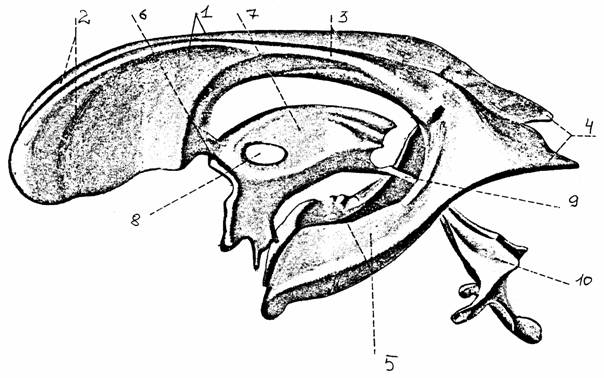
Рис. 28.
Желудочки мозга [15]:
1 - боковые
желудочки, 2 - передний рог, 3 - центральная часть, 4 -
задний
рог, 5 -
нижний рог, 6 - межжелудочковое отверстие, 7 - III
желудочек,
межталамическое сращение, 9 - водопровод,
10 - IY желудочек
Боковые, латеральные
желудочки мозга (правый и левый) лежат внутри обоих полушарий большого
мозга. В каждом из боковых желудочков различают части: передний, задний и нижний рога, центральная
часть. Каждая из них соответствует одной из долей полушария большого мозга:
передний рог залегает в лобной доле, задний - в затылочной, нижний - в височной,
центральная часть - в теменной. Третий
желудочек мозга располагается в срединной сагиттальной плоскости и
сообщается с боковыми желудочками посредством впереди лежащих межжелудочковых
отверстий, с четвертым желудочком -
посредством водопровода мозга. Боковые стенки третьего желудочка образованы
медиальными поверхностями зрительных бугров. Четвертый желудочек внизу
сообщается с полостью спинного мозга его спинномозговым каналом. В задних
отделах четвертого желудочка находятся два отверстия, через которые полость
желудочка сообщается с субарахноидальным пространством, - боковые отверстия четвертого желудочка
(отверстия Люшка). В самой каудальной части
четвертого желудочка есть участок, где его крыша сильно истончается и в ней
образуется отверстие, через которое может выходить часть спинномозговой жидкости
(срединное отверстие четвертого
желудочка, или отверстие Мажанди).
В желудочках мозга находится сосудистое сплетение, выполняющее
функцию секреции цереброспинальной жидкости. Большая его часть равномерно
распределена между четвертым и латеральными желудочками. Около 10% сосудистого
сплетения расположено в третьем желудочке. Во взрослом мозге на долю сосудистого
сплетения приходится, как правило, не более 0,25% массы всего мозга (у человека
2 - 3 г).
8. КРОВЕНОСНЫЕ СОСУДЫ
МОЗГА
Артериальное кровоснабжение осуществляется из двух источников: передние отделы мозга снабжаются кровью через внутренние сонные артерии, а задние - через две позвоночные артерии, которые, сливаясь у основания продолговатого мозга, образуют основную артерию, которая в свою очередь заканчивается двумя задними мозговыми артериями. Системы внутренних сонных и основной артерий объединяются посредством двух задних соединительных артерий на нижней поверхности мозга. Каждая внутренняя сонная артерия делится на три основные ветви: переднюю, среднюю артерии большого мозга (ее проксимальный ствол - задняя соединительная артерия), а также переднюю артерию сосудистого сплетения. Обе передние артерии соединяются посредством передней соединительной артерии, завершающей формирование артериального круга большого мозга. От каждого из трех основных сосудов отходят перфорирующие артерии, идущие к глубоким подкорковым ядрам, а затем образующие сеть, которая дает начало другим перфорирующим артериям, снабжающим кору мозга. Мелкие ветви перфорирующих артерий выполняют в отношении отдельных нейронных групп функцию “конечных артерий”: закупорка одного из таких сосудов приводит к гибели нейронов в снабжаемой им области. Однако закупорка по крайней мере некоторых ветвей поверхностной артериальной сети не всегда сопровождается серьезными корковыми поражениями в силу хорошего развития системы коллатеральных анастомозов.
Артериальное кровоснабжение спинного мозга осуществляется передней и задней спинномозговыми артериями, которые отходят от позвоночной артерии. Отходящие от них перфорирующие сосуды снабжают кровью серое вещество мозга.
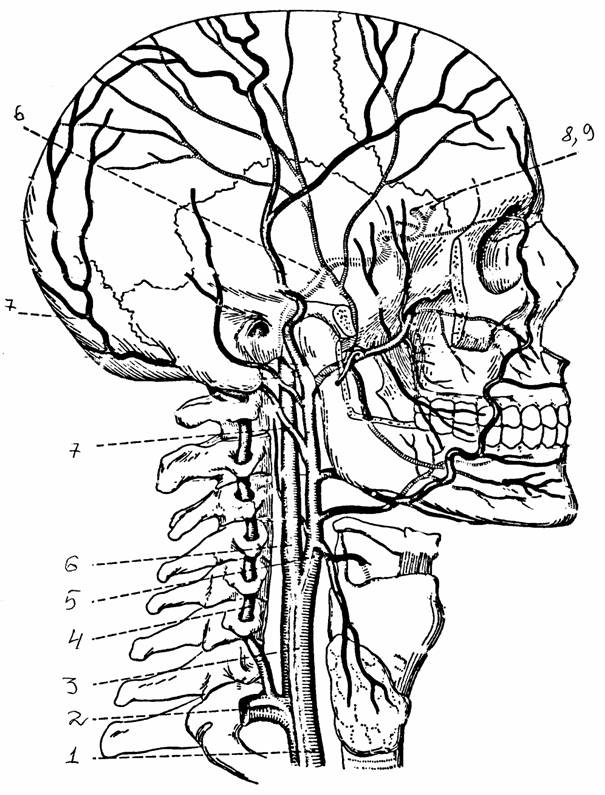
Рис. 29.
Кровоснабжение головного мозга [15]:
1 -
плечеголовной ствол; артерии: 2 - подключичная,
3 - общая сонная,
4 -
позвоночная, 5 - наружная сонная, 6 - внутренняя сонная, 7 -
затылочная,
8, 9 -
средняя и передняя мозговые
|
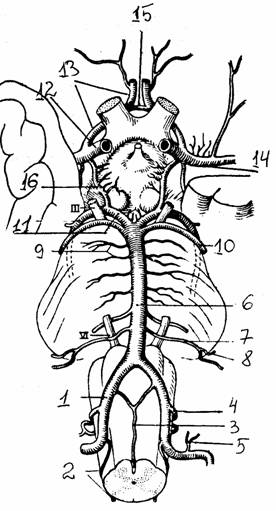
Рис.
30. Артерии основания мозга [6]: Ш,
IY - нервы; артерии: 1 -
позвоночная, 2 - задняя спинномозговая, 3 -
передняя спинномозговая, 4 - задняя нижняя мозжечковая, 5 -
передняя менингиальная ветвь, 6 -
базилярная, 7 -
передняя нижняя мозжечковая, 8 -
лабиринта, 9 - моста, 10 - верхняя мозжечковая, 11 - задняя
мозговая, 12 -
внутренняя сонная, 13 - передняя мозговая, 14 - средняя
мозговая, 15 -
передняя соединительная, 16 -
задняя соединительная |
Венозный отток от передних отделов
мозга идет в систему поверхностных и глубоких вен. Поверхностная система состоит
из вен, отводящих кровь от коры головного мозга и вливающихся в синусы твердой
мозговой оболочки: верхний сагиттальный,
поперечный и пещеристый. Система
глубокого венозного оттока состоит из двух групп сосудов: вены, расположенные
выше и ниже промежуточного мозга и базальных ядер. Вены расположенные более
дорсально, соединяются в парные внутренние вены большого мозга. Под
валиком мозолистого тела эти вены сливаются, образуя большую вену мозга (вена Галена),
которая вливается в прямой синус.
Вены основания мозга сливаются в основную
вену. По ходу в нее вливаются вены, отводящие кровь от промежуточного мозга
и области покрышки среднего мозга.
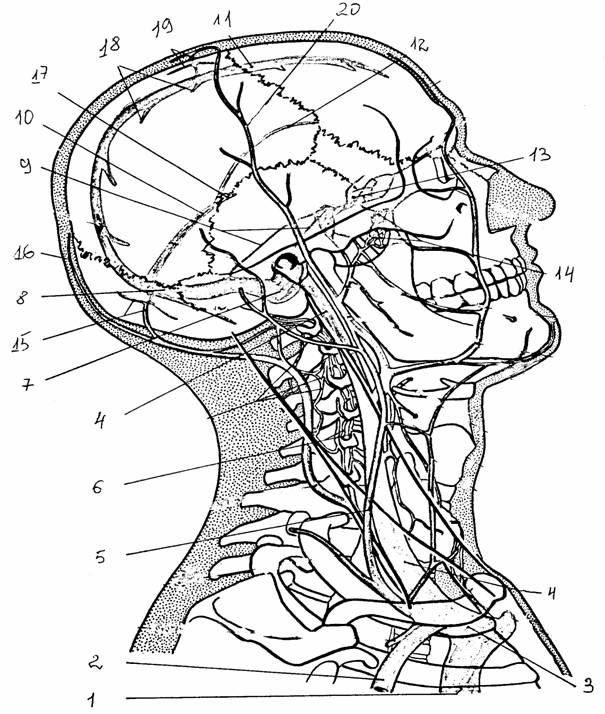
Рис. 31.
Вены головы и синусы [15]:
вены: 1 -
верхняя полая, 2 - подключичная, 3 - плечеголовная,
4 -
внутренняя яремная, 5 - наружная яремная, 6 - позвоночная;
синусы:
7 -
сигмовидный, 8 - поперечный, 9 - каменистые верхний,
нижний,
10 -
прямой, 11 - верхний сагиттальный, 12 - нижний
сагиттальный,
13 -
пещеристый, 14 - крыловидное сплетение; вены: 15 -
затылочные
диплоическая и эмиссарная, 16 - затылочная, 17 - большая
мозговая,
18 -
верхние мозговые, 19 - теменная эмиссарная, 20 -
височная поверхностная
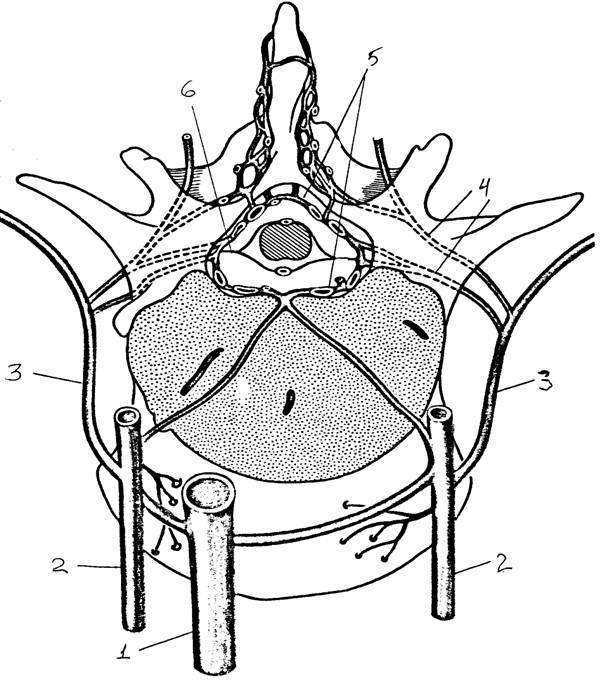
Рис. 32.
Венозное кровоснабжение позвоночного столба (цит. по [14]):
вены: 1 -
нижняя полая, 2 - восходящая поясничная, 3 - поясничная,
4 -
межпозвоночные;
5 - переднее,
заднее внутренние позвоночные
венозные
сплетения, 6 - корешковые вены
Венозный отток от спинного мозга осуществляется через передний продольный венозный ствол, а также через множество задних продольных вен. Все эти сосуды связаны друг с другом хорошо развитой системой анастомозов, образующих сплетение по всей поверхности спинного мозга.
9. ОНТОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ
ЧЕЛОВЕКА
Морфогенез
Нервная система человеческого эмбриона, формирующаяся из эктодермы,
представлена двумя структурами: первичной
полоской и нотохордом. Первичная полоска является
утолщением эктодермы эмбриона, представляющего к концу третьей недели овальный
диск около 1,5 см в длину. Рострально ее находится
другое утолщение эктодермы - первичный
(гензеновский) узелок. Узелок, втягиваясь,
образует бластопор. В ростральном направлении между эктодермой и энтодермой из
первичного узелка мигрирует тяж клеток, образующий нотохорд. При дальнейшем втягивании бластопора, переходящего
в нотохорд, образуется нотохордальный канал. Эктодерма, покрывающая нотохорд, утолщается, формируя нервную пластинку.
В дальнейшем нервная пластинка прогибается с образованием сначала нервной бороздки, а в конце - длинной
полой трубки, лежащей непосредственно
под поверхностью эктодермы, от которой она отделяется. Первой замыкается та
часть нервной трубки, которая образует задний мозг. При замыкании бороздки в
каудальном направлении формируется часть нервной трубки, которая соединяется со
спинным мозгом. Замыкание нервной бороздки в ростральном направлении происходит
одновременно с формированием сегментов спинного мозга. Оно сначала ведет к
образованию среднего, а затем переднего мозга. Последним замыкается небольшое
отверстие на мозговом конце, называемое передним нейропором. По мере замыкания нервной трубки эктодермальные клетки боковых краев нервной бороздки
выталкиваются в сторону, образуя продолговатый тяж клеток по обеим сторонам
трубки - нервный гребень. Процесс
формирования нервной трубки называется нейруляцией.
По завершении нейруляции клетки нервного гребня мигрируют наружу и
дают начало спинальным ганглиям, периферическим ганглионарным нейронам симпатической нервной системы, шванновским клеткам, клеткам спинальных ганглиев, а также
клеткам, образующим внутренние листки оболочек мозга. Сформировавшись, длинная
полая нервная трубка подвергается дальнейшим изменениям. На ранней стадии она
подразделяется на длинную каудальную трубку, образующую спинной мозг, и более
широкие ростральные сегменты, которые превращаются в головной мозг.
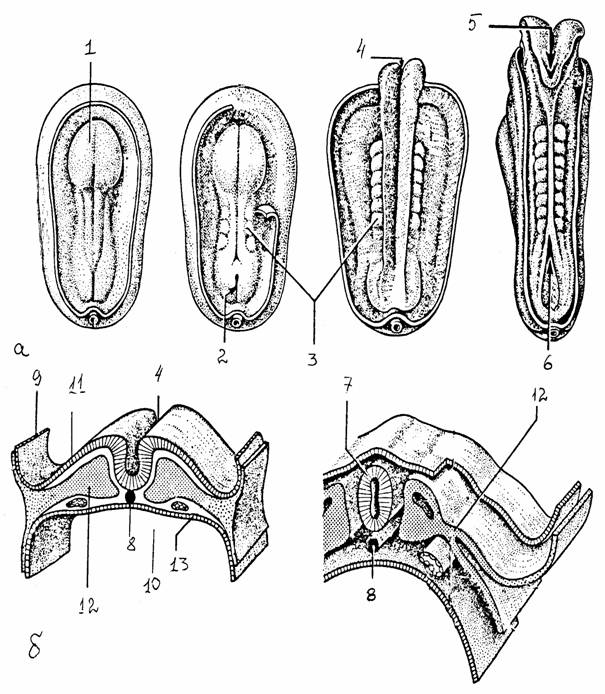
Рис. 33.
Дифференцировка зародышевых листков [11]:
а -
зародышевый диск на желточном мешке,
б - схема
поперечного разреза зародышевого диска;
1 - нервная
пластинка, 2 - первичная бороздка, 3 - сомиты, 4 - нервная бороздка,
5, 6 -
передний, задний нейропор, 7 - нервная трубка, 8 -
нотохорд, 9 - амнион,
10 -
полость желточного мешка, 11 - эктодерма, 12 - мезодерма, 13 -
энтодерма
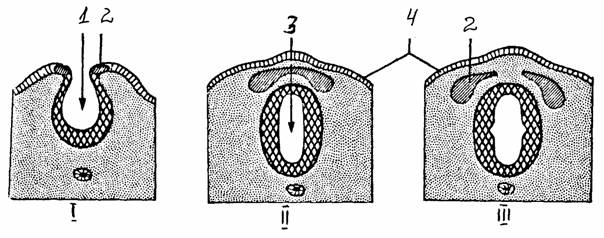
Рис. 34.
Схема образования нервной трубки и нервного гребня [11]:
I - образование нервной
бороздки, ее погружение, II - образование
нервной
трубки,
нервного гребня, Ш - миграция клеток нервного гребня;
1 - нервная
бороздка, 2 - нервный гребень, 3 - нервная трубка, 4 -
эктодерма
Головной конец вскоре подразделяется на три расширения, первичные мозговые пузыри. Полости этих пузырей сохраняются в мозге взрослого в видоизмененной форме и образуют полости желудочков и сильвиева водопровода.
Самым ростральным отделом нервной трубки является передний мозг, за ним следует средний мозг, и третий - наиболее каудальный сегмент - образует ромбовидный мозг. В последующем развитии передний мозг делится на конечный мозг (полушария большого мозга и некоторые базальные ядра) и промежуточный мозг. С каждой стороны промежуточного мозга вырастает глазной пузырь, формирующий нервные элементы глаза. Средний мозг сохраняется как единое целое. Ромбовидный мозг подразделяется на продолговатый мозг, мозжечок и мост.
Гистогенез
Первичная нервная трубка в спинальном отделе рано делится на три
слоя: 1) самый внутренний эпендимный слой, содержащий зачатковые клетки; 2)
промежуточная зона (мантийный слой),
куда мигрируют пролиферирующие клетки; 3) наружный краевой слой, который в зрелом
мозге содержит миелинизированные волокна. Эпендимный (герменативный) слой дает начало нервным и глиальным элементам ЦНС. Каждая зачатковая клетка делится на
две дочерние, одна из которых мигрирует в
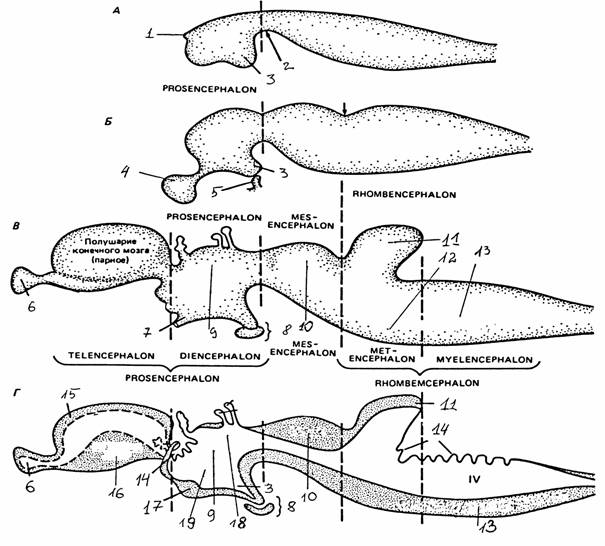
Рис. 35.
Схема последовательности
развития
основных отделов и структур мозга [3]:
А - в
нервной трубке дифференцированным является только
первичный
передний
мозг (prosencephalon); Б - намечаются три
основных отдела
(стадия
трех мозговых пузырей); В, Г - развиты пять отделов
головного
мозга (стадия пяти мозговых пузырей);
Prosencephalon -
первичный
передний мозг,
Rhombencephalon
- первичный
задний мозг, Mesencephalon -
средний мозг,
Telencephalon - конечный мозг,
Diencephalon -
промежуточный
мозг,
Metencephalon -
задний мозг,
Myelencephalon - продолговатый
мозг;
1 - нейропор, 2 - вентральная борозда, 3 - воронка, 4 - глазной
пузырь,
5 -
гипофизарный карман, 6 - обонятельная луковица, 7 - глазной
стебелек,
8 -
гипофиз, 9 - таламус, 10 - крыша, 11 - мозжечок, 12 -
мост,
13 -
продолговатый мозг, 14 - сосудистое сплетение, 15 - кора
(плащ),
16 -
базальные ядра, 17 - зрительный перекрест,
18 - эпиталамус, 19 - гипоталамус
мантийный слой, а вторая остается в герменативном. Мигрирующие в мантийный слой клетки превращаются в нейробласты, которые в дальнейшем дифференцируются в зрелые нейроны. Клетки, остающиеся в глубине эпендимного слоя, превращаются в астроцитобласты. Некоторые клетки, оставаясь недифференцированными, отделяются от эпендимного слоя (медуллобласты) и образуют олигодендроциты.
Таблица
2
Основные
изменения в процессе развития нервной системы человека
|
Возраст зародыша, недели |
Длина зародыша, мм |
Развитие
мозга |
|
2,5 |
1,5 |
Намечается
нервная бороздка |
|
3,5 |
2,5 |
Нервная
бороздка закрывается. Нервный гребень имеет вид сплошной
ленты |
|
4,0 |
5,0 |
Нервная
трубка замкнута. Образовались три первичных
мозговых пузыря. Формируются нервы и
ганглии. Сформировались эпендимный,
мантийный и
краевой слои |
|
5,0 |
8,0 |
Формируются
5 мозговых пузырей. Намечаются полушария большого
мозга |
|
6,0 |
12,0 |
Образуются
три первичных изгиба нервной трубки. Нервные сплетения сформированы.
Симпатические узлы формируют сегментарные скопления. Намечаются мозговые
оболочки |
|
7,0 |
17,0 |
Полушария
мозга достигают большого размера. Появляются сосудистые сплетения. Хорошо
выражены полосатое тело и зрительный бугор |
|
8,0 |
23,0 |
В
коре мозга появляются типичные нервные клетки. Выражены оболочки
мозга |
|
10,0 |
40,0 |
Формируется
внутренняя структура спинного мозга |
|
12,0 |
56,0 |
Формируются
общие структурные черты головного мозга. Начинается дифференцировка клеток
нейроглии |
|
16,0 |
112,0 |
Полушария
покрывают большую часть мозгового ствола. Различаются доли головного
мозга. Появляются бугры четверохолмия. Выражен
мозжечок |
|
20-40 |
160-520 |
Завершается
формирование комиссур мозга (20 нед.).
Начинается миелинизация спинного мозга.
Появляются типичные слои коры головного мозга (25 нед.). Формируются борозды и извилины головного мозга
(28-30 нед.). Начинается миелинизация головного мозга (36-40 нед.) |
10. АНАТОМИЯ СЕНСОРНЫХ СИСТЕМ
ПОЗВОНОЧНЫХ
Зрительная сенсорная
система
Глаз позвоночных устроен по камерному типу и работает по принципу создания изображения путем рефракции. Светопреломляющий аппарат образован роговицей и хрусталиком. Дно глазного бокала выстилает сетчатка, пространство между ней и хрусталиком заполнено стекловидным телом. Снаружи глазное яблоко покрыто непрозрачной волокнистой тканью - склерой, которая на переднем полюсе глаза переходит в прозрачную роговицу. Хрусталик делит глазное яблоко на переднюю камеру, заполненную жидкостью, и камеру большего размера, заполненную стекловидным телом. Изнутри к склере прилегает сосудистая оболочка, богатая кровеносными сосудами. Ее продолжением являются ресничное тело и радужка. Сокращение или расслабление волокон ресничного тела приводит к изменению натяжения цинновых связок, ответственных за изменение кривизны хрусталика. Радужка выполняет роль диафрагмы: диаметр отверстия в ней (зрачок) изменяется за счет работы кольцевых и радиальных гладких мышц.
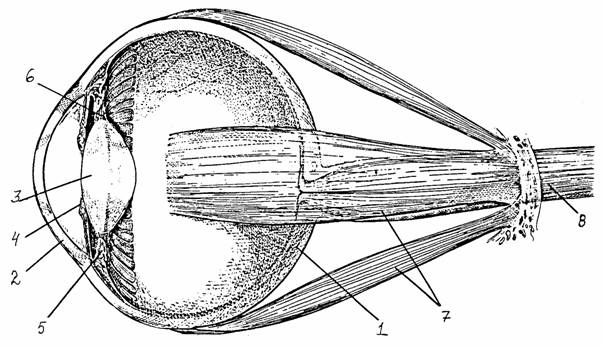
Рис. 36.
Схема строения глазного яблока [7]:
1 - склера,
2 - роговица, 3 - хрусталик, 4 - радужная оболочка, 5 - ресничная мышца, 6 -
цинновы связки, 7 - наружная мышца глаза, 8 -
зрительный нерв
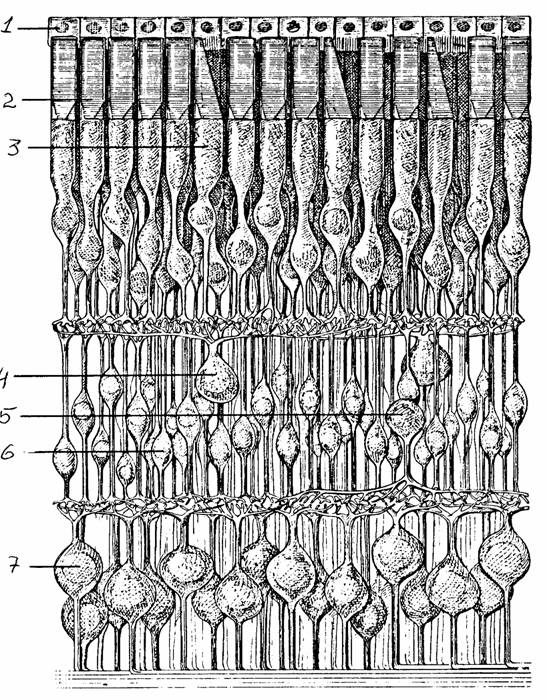
Рис. 37.
Поперечный разрез сетчатки [7]:
1 -
пигментная клетка, 2 - палочка, 3 - колбочка, 4 - горизонтальная
клетка,
5 - амакриновая клетка, 6 - биполярная клетка, 7 - ганглиозная клетка
Самый внутренний слой глаза - сетчатка - состоит из фоторецепторов (колбочек и палочек), четырех типов нервных клеток, глиальных клеток (мюллеровские клетки) и пигментных клеток.. Сетчатка позвоночных имеет инвертированную структуру: фоторецепторы располагаются в самом удаленном от стекловидного тела слое. В глазу человека морфологически различаются два класса фоторецепторов: палочки (около 120 млн.) и колбочки (около 6 млн.). Фоторецепторы соединены синаптическими контактами с биполярными и горизонтальными клетками. Биполяры передают сигналы от фоторецепторов в слой амакриновых и ганглиозных клеток. Аксоны ганглиозных клеток, граничащих со стекловидным телом, направляются к оптическому диску, или слепому пятну, где они собираются вместе, проходят через склеру и образуют зрительный нерв. В месте пересечения сетчатки оптической осью находится углубление (фовеа), которое содержит из фоторецепторов только колбочки.
Волокна зрительного нерва проецируются в ядра верхних бугров четверохолмия (средний мозг), но в большей степени в латеральное коленчатое тело (ЛКТ). Стволы зрительных нервов образуют перекрест (хиазму), в котором происходит частичное перекрещивание волокон. Аксоны ганглиозных клеток от наружной (височной) половины сетчатки попадают в ипсилатеральное коленчатое тело, от внутренней - в контрлатеральное. В ЛКТ входные сигналы от ганглиозных клеток различных глаз проецируются в разные слои: контрлатеральные волокна попадают в 1-й, 4-й и 6-й слои, а ипсилатеральные - во 2-й, 3-й и 5-й. Аксоны нейронов ЛКТ идут в 17-е поле (по Бродману) в затылочной области коры - первичное зрительное поле. Кроме того, зрительная информация поступает также в 18-е и 19-е поле.
Слуховая сенсорная система
млекопитающих
Наружное ухо включает ушную раковину и наружный слуховой проход. Среднее ухо содержит косточки (молоточек, наковальню и стремечко), связанные с одной стороны с барабанной перепонкой, а с другой стороны - с овальным окном внутреннего уха. В среднем ухе есть тимпанальная мышца и мышца стремечка, регулирующие амплитуду колебаний слуховых косточек. Внутреннее ухо (улитка) состоит из трех параллельных трубчатых каналов (лестниц), расположенных в спирально закрученном (у человека 2,5 витка) костном канале: вестибулярная, средняя и барабанная лестница. Вестибулярная и барабанная лестницы соединяются у вершины улитки (геликотрема) и заполнены перилимфой. У основания улитки оба эти канала отделены от полости среднего уха сходными структурами. Овальное окно, ведущее в вестибулярную лестницу, замкнуто стремечком, а круглое окно в конце барабанной лестницы затянуто тонкой перепонкой, которая отделяет ее от полости среднего уха. Средняя лестница заполнена эндолимфой. Границу между вестибулярной и средней лестницей образует вестибулярная (рейснерова) мембрана, а между средней и барабанной лестницей - базилярная (основная) мембрана, на которой находится собственно сенсорный аппарат - кортиев орган.
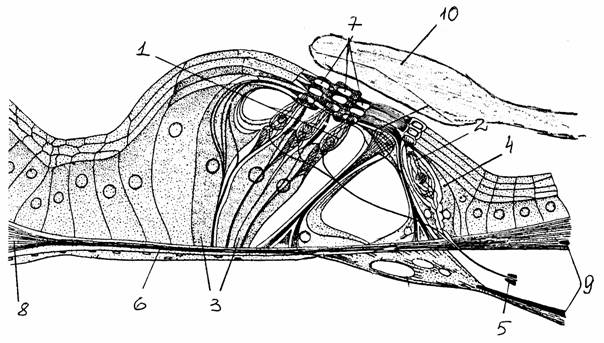
Рис. 38.
Кортиев орган (с изменениями по
[15]):
1, 2 -
внешние и внутренние волосковые клетки, 3, 4 - внешние
и
внутренние поддерживающие (опорные) клетки, 5 - нервные
волокна,
6 -
базилярная мембрана, 7 - отверстия ретикулярной (сетчатой)
мембраны,
8 -
спиральная связка, 9 - костная спиральная пластинка,
10 - текториальная (покровная) мембрана
В кортиевом органе находятся волосковые клетки, окруженные поддерживающими клетками. Волосковые клетки несут на своей поверхности выросты - стереоцилии. Различают внутренние (около 3500 у человека) и наружные (около 20 000) волосковые клетки. Наружные расположены тремя рядами, внутренние образуют один. Над кортиевым органом лежит желатинообразная масса, текториальная (покровная) мембрана. Она прикреплена к внутренней стороне улитки и прикасается к стереоцилиям волосковых клеток. Волосковые клетки кортиева органа не имеют аксонов. Тела нервных клеток, передающих возбуждение от этого органа к ЦНС, находятся в спиральном ганглии, который лежит в улитке, завиваясь вокруг нее вместе с каналами. Нервные клетки этого ганглия биполярные: один отросток идет на периферию к волосковым клеткам, другой - в составе слухового нерва к ЦНС. 95% волокон слухового нерва (у человека их всего около 25 000) связаны только с внутренними волосковыми клетками.
Первичные афферентные волокна нейронов спирального ганглия идут к кохлеарному ядру. От его вентральной части сигналы проводятся к ипси- и контрлатеральным верхне-оливарным комплексам. Аксоны нейронов дорсального кохлеарного ядра переходят на противоположную сторону и образуют синапсы с нейронами ядра латерального лемниска. Аксоны последних частично переходят на противоположную сторону и идут через два ядра переключения - нижние бугры четверохолмия и медиальное коленчатое тело (МКТ). Нейроны МКТ посылают аксоны к первичной слуховой коре в височной доле.
Гравитационная сенсорная система
млекопитающих
Вестибулярный аппарат непосредственно
связан с улиткой внутреннего уха (вместе они образуют лабиринт) и содержит отолитовый
орган и полукружные каналы.
Отолитовый орган разделен на два мешочка - саккулюс и утрикулюс, и рецепторные клетки группируются в макуле каждого из них. Каждый полукружный канал имеет расширение (ампулу), внутри которого рецепторы группируются в гребешки. Каждая сенсорная клетка несет на своей поверхности киноцилию (ресничка, содержащая 9 пар микротрубочек) и какое-то количество стереоцилий. В макулах поверх волосков лежат отолиты. Они в основном состоят из кристаллов карбоната кальция (отокониев), склеенных вместе желеобразным веществом в толстую отолитовую мембрану. В гребешках волоски входят в желеобразный свод, который простирается до другой стороны ампулы.
К волосковым клеткам подходят афферентные волокна нейронов, находящихся в вестибулярном ганглии, лежащем в височной кости близ вестибулярного органа. Аксоны, по которым эти клетки передают сигналы в ЦНС, входят в состав VIII нерва и заканчиваются на нейронах ядер вестибулярного комплекса продолговатого мозга: верхнее (ядро Бехтерева), латеральное (ядро Дейтерса), медиальное (ядро Швальбе) и нижнее ядра. Вестибулярные ядра связаны со всеми отделами ЦНС: спинным мозгом, мозжечком, ретикулярной формацией, глазодвигательными ядрами, корой головного мозга, вегетативной нервной системой. Можно выделить три главные проекционные системы: вестибулоспинальная, вестибуломозжечковая и вестибулоокулярная.
Соматическая сенсорная
система
В коже располагается ряд своеобразных рецепторных образований,
обеспечивающих механическую, температурную и болевую чувствительность.
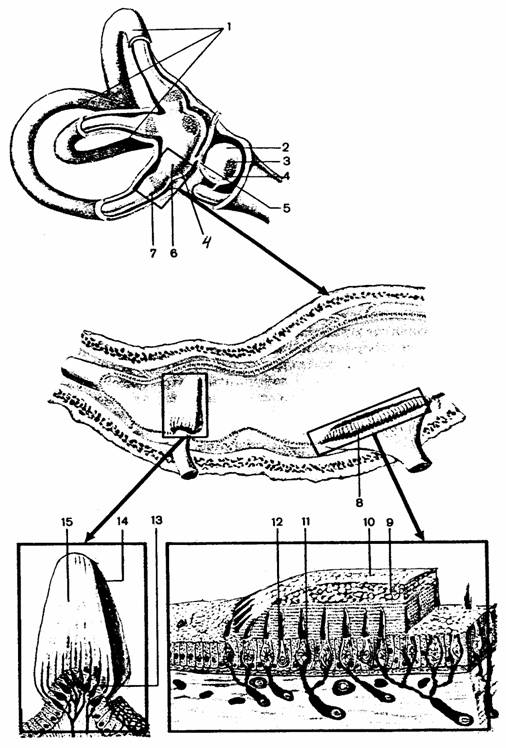
Рис. 39.
Орган равновесия [4]:
1 -
полукружные каналы, 2 - преддверие, 3 - сферический мешочек (саккулюс),
4 - пятно
(макула), 5 - эндолимфатический проток, 6 - эллиптический
мешочек
(утрикулюс), 7 - ампула, 8 - отолитовый аппарат, 9 - статоконии, 10 - мембрана
статоконий, 11 - поддерживающие
клетки, 12, 13 - волосковые сенсорные
клетки, 14
- ампульный гребешок, 15 - купол
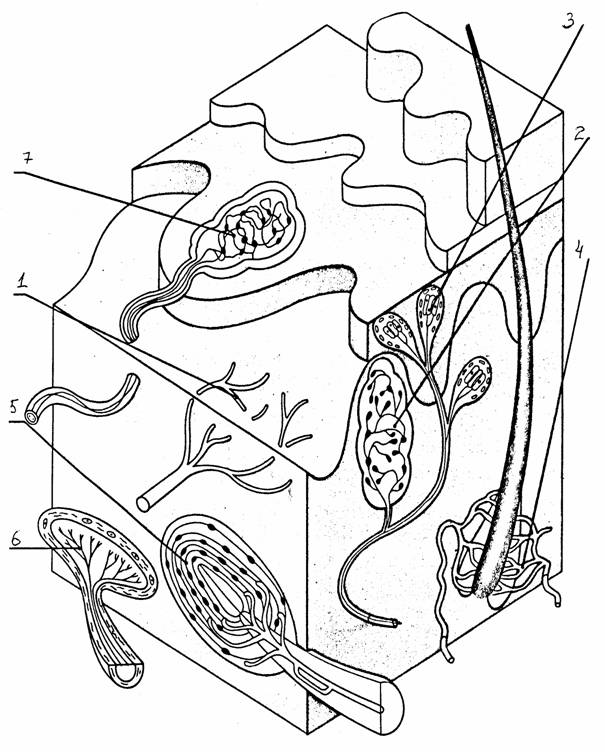
Рис. 40.
Типы соматосенсорных рецепторов [10]:
1 -
свободные нервные окончания, 2 - тельце Мейснера, 3 -
диск Меркеля,
4 -
рецепторы волосяного фолликула, 5 - тельца Паччини,
6 -
окончания Руффини, 7 - колбы Краузе
Наиболее простой тип - свободные нервные окончания. Кроме них встречаются морфологически дифференцированные образования. У человека в сосочковом слое кожи без волосяного покрова имеются конусовидные покрытые капсулой тельца Мейснера и диски Меркеля. В волосистых участках кожи располагаются рецепторы волосяного фолликула и окончания Руффини. В более глубоких слоях кожи и подкожной клетчатки находятся тельца Паччини, состоящие из многослойной наружной капсулы, внутренней колбы и заключенной в нее части афферентного нервного волокна, лишенного миелина.
Тела нейронов, афферентные аксоны которых подходят к кожным
рецепторам или сами выполняют рецепторную функцию, находятся в спинальных
ганглиях и передают в головной мозг информацию по двум основным путям: лемнисковому (пучки Голля и Бурдаха) и спинно-таламическому. Центральные зоны этой
сенсорной системы находятся у человека в области постцентральной извилины (первая соматосенсорная область) и
областях, примыкающих к слуховой коре (вторая соматосенсорная
область)
Проприоцептивная сенсорная система
Специфические рецепторы скелетно-мышечной системы позвоночных - мышечные веретена и сухожильные органы Гольджи. Мышечные веретена находятся в составе скелетных мышц. Их основу составляет пучок интрафузальных мышечных волокон, имеющих афферентную и эфферентную иннервации. Сухожильные органы располагаются в зоне соединения мышечных волокон с сухожилием. Оба типа рецепторов иннервируются афферентными аксонами нейронов спинномозговых ганглиев. В кору головного мозга (соматосенсорная область) информация от проприоцепторов поступает в составе пучков Голля и Бурдаха.
Хеморецепторные сенсорные
системы
Вкусовая система. Язык
человека покрыт слизистой оболочкой, складки которой образуют сосочки (грибовидные, желобоватые, листовидные), на поверхности сосочков
находятся вкусовые почки. Отдельная
вкусовая почка имеет около 70 мкм в высоту и около 40 мкм в диаметре. Всего у
человека около 2000 вкусовых почек, каждая из которых содержит 40-60 отдельных
клеток трех типов: сенсорные, опорные и
базальные. Вкусовые сенсорные клетки лишены аксонов, информация передается
от них через синапсы на окончания сенсорных волокон на вкусовой почке. Эти
волокна
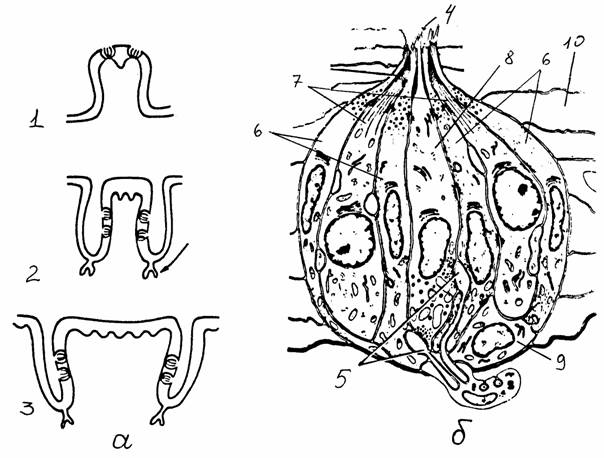
Рис. 41.
Типы вкусовых сосочков (а), строение вкусовой почки (б) (цит. по [12]):
вкусовые
сосочки: 1 - грибовидные, 2 -
листовидные, 3 - желобоватые;
4 -
вкусовая пора, 5 - синапсы, 6, 7 - поддерживающие (опорные)
клетки,
9 -
базальные клетки, 10 - эпителиальные клетки
отходят от ганглиозных клеток черепномозговых нервов - ветви VII (барабанная струна) и IX. Эти нейроны образуют центральные синапсы в продолговатом мозгу - на нейронах ядра одиночного тракта. В свою очередь нейроны ядра образуют синапсы на нейронах таламических ядер, а те посылают аксоны в постцентральную извилину коры больших полушарий.
Обонятельная система. У
человека обонятельные рецепторы сосредоточены на небольшом участке эпителия
верхней и (частично) нижней носовой раковины (обонятельный эпителий). Кроме
обонятельных рецепторов в этом эпителии располагаются базальные клетки. На своем
апикальном полюсе обонятельные клетки несут видоизмененные реснички, погруженные
в слой слизи, которая покрывает обонятельный эпителий. Аксоны обонятельных
клеток образуют толстые пучки под сенсорным эпителием (обонятельные волокна), которые идут к
обонятельным луковицам, проходя через
решетчатую кость. Клеточные элементы обонятельной луковицы расположены
слоями (по направлению к центру): 1) слой filia olfactoria; 2) слой гломерул; 3)
наружный плексиморфный слой; 4) слой тел митральных
клеток; 5) слой клеток-зерен. Весьма крупные митральные клетки отдают главный
дендрит, ветвления которого образуют синапсы с волокнами обонятельных клеток. На
каждой митральной клетке образуют синапсы до 1000 волокон. Аксоны митральных
клеток образуют обонятельный тракт, передающий сигналы во многие области мозга,
в том числе в обонятельную луковицу противоположной стороны, обонятельную кору,
гиппокамп и через миндалину к ядрам гипоталамуса. К
обонятельной луковице подходит множество эфферентных волокон от головного мозга:
от обонятельных корковых областей, базальной части конечного мозга и среднего
мозга.
СПИСОК
ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Зайцева О.В. Распределение
афферентных элементов в центральной нервной системе обыкновенного прудовика
//
Архив анатомии,
эмбриологии и гистологии. 1981. Т. 81. № 11.С. 35-42.
2. Иванов А.В., Мончадский А.С., Полянский Ю.И., Стрелков А.А. Большой
практикум по зоологии беспозвоночных. Ч. 2. М.: Высшая школа,1983. 543
с.
3. Ромер А., Парсонс Т. Анатомия
позвоночных. Т. 2. М.: Мир, 1992. 406 с.
4. Сапин М.Р.,Билич Г.Л. Анатомия
человека. Т 2. М.: Высшая школа, 1996. 432 с.
5. Свидерский В.Л. Основы нейрофизиологии насекомых. Л.: Наука.
Ленинградское отд-ние, 1980. 280
с.
6. Фениш Х., Даубер В. Карманный
атлас анатомии человека. Минск: Вышэйшая
школа,1996.
464 с.
7. Хьюбел Д. Глаз, мозг, зрение. М: Мир,1990. 240
с.
8.
Biologiekolleg.
Berlin: Cornelsen-Velhagen&Klasing, 1981. 464
S.
9.
Eckert
R. Tierphysiologie. Stuttgart,New York: Georg Thieme Vrlag,1986. 698 S.
10.
Der
Mensch. Bonn: Verlag des
Borromaeusvereins,1986 208
S.
11.
Schumacher
G.-H. Embryonale Entwicklung
des Menschen. Berlin: VEB Verlag Volk und Gesundheit, 1987.
282 S.
12.
Shepherd
G.M. Neurobiologie. Berlin,
Heidelberg, New York: Springer-Verlag, 1993. 624
S.
13.
Ude
J., Koch M. Die Zelle: Atlas
der Ultrastruktur. Jena,
Stuttgart: Gustav Fischer Verlag, 1994. 312
S.
14.
Waldeyer
A. Anatomie des Menschen.
Teil 1. Berlin, New York: de Gruyter, 1987. 494 S.
15.
Waldeyer
A. Anatomie des Menschen.
Teil 2. Berlin, New York: de Gruyter, 1986. 638 S.